La Collection Ouellet-Robert possède un magnifique ensemble de spécimens de libellules et demoiselles (ordre des Odonates) grâce en grande partie aux efforts de son fondateur éponyme, Adrien Robert. En effet, la collection d’Odonates est si bonne que nous avons choisi une demoiselle, le caloptéryx élancé (Calopteryx amata), comme emblème de la collection.
Calopteryx amata, mâle
Grâce au financement de démarrage de Canadensys de la Fondation canadienne pour l’innovation, la numérisation des données des spécimens d’Odonates de la collection a été faite avant 2012. Ensuite, pour mieux intégrer les activités des différentes collections d’insectes au Québec, et peut-être pour fédérer une collaboration plus ambitieuse, avec du financement du Centre de la sciences de la biodiversité du Québec, des collègues entomologistes ont visité six autres collections du Québec et ajouté leurs données de spécimens Odonata au mélange. Le résultat est un ensemble de données impressionnant, comprenant 37 000 enregistrements d’occurrence, provenant de 616 emplacements différents, pour 137 espèces. Cet ensemble de données, dont la description est publiée dans le Biodiversity Data Journal, est gratuit à télécharger et à utiliser pour la modélisation de la distribution des espèces, l’analyse des changements dans le temps, et une gamme d’autres sujets de recherche. Étant donné que de nombreuses espèces d’Odonates au Québec se trouvent aux limites nord de leur aire de répartition, leurs expansions possibles pourraient être de bons indicateurs pour mesurer les effets des changements climatiques.
Calopteryx amata, femelle
Cette publication est la deuxième, en moins d’un an, qui met l’accent sur les activités de la Collection Ouellet-Robert. La première était un aperçu de trois initiatives d’informatisation, y compris la numérisation des Odonates, tandis que cette deuxième se concentre sur les données elles-mêmes.
Texte et figures (sauf si indiqué) CC BY-SA 2018, par les auteures.
Anax junius (Drury, 1773) l’anax précoce
L’espèce Anax junius a été décrite pour la première fois en 1773 par Drury. Notre spécimen (Figure 1) a été attrapé le 06 septembre 2018 à l’aide d’un filet fauchoir sur le Chemin du Lac Croche menant à la station de biologie des Laurentides (Figure 2) aux coordonnées suivantes: Latitude 45.9842 | Longitude -74.009
Figure 1 : Vue dorsale de Anax junius (Photo prise par Lucille Joyeux)
Figure 2 : Chemin du Lac Croche (Photo prise par Zachary Bélisle)
Classification
Embranchement Arthropoda
Sous-embranchement Hexapoda
Classe Insecta
Ordre Odonata
Sous-ordre Anisoptera
Famille Aeshnidae
Genre Anax
Espèce Anax junius
Identification
Pour identifier précisément notre spécimen, nous avons utilisé deux clés d’identification (Robert, 1957 ; Pilon et Lagacé, 1998). Le processus d’identification de notre spécimen a été relativement simple grâce à sa grande taille (7.4cm) qui a facilité nos observations.
Nous avons d’abord chercher la famille de notre odonate grâce à la clé d’identification des familles de Robert A. (1957).
On remarque que la largeur de la base des ailes est différente, en effet, la base de l’aile postérieure est plus large que la base de l’aile antérieure. Ce caractère nous a permis d’approuver que notre spécimen provient du sous-ordre Anisoptera, soit plus communément des libellules. Ensuite, on voit que la distance entre l’arculus et le triangle est à peu près égale dans les deux paires d’ailes (Figure 3). Le ptérostigma, qui est la tâche noire à l’extrémité de chaque aile, est pourvu d’une grosse nervure oblique à son bord interne, et on observe également que les yeux se touchent. Ces 3 observations de caractères permettent de dire que notre anisoptère est plus précisément de la famille des Aeshnidae.
Figure 3 : Caractéristiques de l’aile antérieure et postérieure de Anax junius (Photo prise et modifiée par Lucille Joyeux)
Une fois notre famille trouvée, nous avons cherché l’espèce grâce à la clé d’identification de Pilon J-G. et Lagacé D. (1998).
Cette recherche a été très simple puisque notre espèce a un caractère bien précis qui la différencie des autres Aeshnides de l’Amérique du Nord : son thorax de couleur vert. Ce caractère est unique à l’espèce Anax junius en Amérique du Nord.
Nous avons également pu déterminer facilement le sexe de notre Anax junius puisque cette espèce a un dimorphisme sexuel visuel : la couleur de son abdomen. Chez les mâles, l’abdomen est de couleur bleue tandis que chez la femelle, l’abdomen est de couleur brune. Notre Anax junius est donc une femelle.
Description
Anax junius fait partie des plus grosses libellules, atteignant 7 ou 8 cm de long. Leurs yeux sont grands, leur thorax est court et robuste et leur abdomen est long et mince. Tous les individus, mâles et femelles, possèdent un thorax vert et une ligne noire qui s’étend sur leur surface supérieure. (Needham et Westfall, 1955). Cependant, les individus sont polymorphes selon le sexe. Les mâles ont le thorax, le visage, et les yeux de couleur verts ternes. Le premier segment de leur abdomen est vert, et les 2 à 6 autres segments sont de couleurs bleu vif après quoi l’abdomen s’estompe en vert terne puis devient sombre. Chez la plupart des femelles, les yeux sont brun, les deux premiers segments abdominaux sont verts, et le reste de l’abdomen est brun rougeâtre (Fraker et Luttbeg, 2012). Chez le juvénile, l’abdomen est vert terne, rougeatre ou brun. Les ailes sont incolores ou orange et prennent une couleur ambrée à mesure qu’elles vieillissent, notamment chez les femelles. (Fraker et Luttbeg, 2012)
Cycle de vie
Les libellules Anax junius sont hémimétaboles, c’est-à-dire sans stade immobile entre la larve et l’adulte.
1. La reproduction a normalement lieu dans un habitat d’eau calme, par exemple les étangs, les lacs, les marais, les marécages, les cours d’eau lents. (Beaton, 2007).
2. Les œufs sont pondus dans des habitats aquatiques et éclosent après au moins 6 ou 7 jours. Les œufs éclosent en nymphes et subissent 9 à 13 instars nymphaux. Les nymphes ont un abdomen ovale allongé, de grands yeux et un grand labium qui peut être étendu pour capturer une proie. Les nymphes Anax junius ont des labiums plats avec des palpes pointues qui sont utilisées pour lancer une proie. Ils sont généralement de couleur marron ou verte. Les derniers stades peuvent atteindre 3,5 à 5 cm de long (Paulson, 2011).
3. Lorsqu’elles atteignent la fin du stade nymphal, les nymphes sortent de l’eau et cherchent un endroit pour émerger de leur exuvie. Leurs ailes se déploient à mesure que les ailes et le corps se dilatent. À ce moment là, elles sont des adultes sexuellement immatures. Elles quittent leur peau nymphale et gagnent rapidement en masse. La cuticule durcit et leur couleur devient plus distinct. Le temps total de développement des nymphes peut prendre presque 1 an. Les nymphes terminent leur développement à l’âge adulte en juin et en juillet. (Hopkins et al, 2011)
4. Les adultes ne vivent que plusieurs semaines à un peu plus d’un mois après avoir atteint l’âge adulte (Paulson, 2011).
Distribution géographique
Anax Junius est répandue majoritairement dans la région du Néarctique. Elles sont présentes sur l’ensemble des États-Unis, du Canada et du Mexique (Figure 4).
Leur aire de répartition s’étend également à Hawaii, à Tahiti, aux Antilles, en Chine et dans la péninsule en Russie. Parfois les individus sont emportés par des tempêtes ou des vents violents dans des régions comme aux Bermudes, au Royaume-Uni et en France. Anax Junius reste l’une des libellules les plus répandues en Amérique du Nord. (Corbet, 1999)
Figure 4 : Distribution géographique de Anax junius selon le IUCN Red List of Threatened Species (Carte prise sur iucnredlist.org le 30 octobre 2018)
Populations de Anax junius
Cette espèce a deux types de population différentes: les populations résidentes et migratrices.
Les populations résidentes restent dans la zone générale d’où elles émergent. Pour les populations résidentes du Nord, les adultes s’accouplent et pondent de fin juillet à août. La progéniture qui en résulte éclot et se développe vers le mi-stade puis hiverne lorsque la température baisse. Le temps total de développement des nymphes peut prendre de 11 à 12 mois pour les résidentes, puisque les nymphes ont terminé leur développement lorsque les températures se réchauffent au printemps, puis apparaissent à l’âge adulte en juin et en juillet (Trottier, 1971).
Les adultes qui migrent vers le Nord tendent à arriver avant l’émergence des résidentes du Nord au printemps. Les adultes migrateurs s’accouplent et pondent en juin. Contrairement aux populations résidentes, le développement nymphal des progénitures migrantes ne prend que 3 à 5 mois et elles ne passent pas l’hiver. Elles émergent généralement en fin août et en septembre. Elles commencent souvent leur migration à l’âge immature, se nourrissant en cours de route pour devenir adultes. Les adultes s’accouplent et pondent pendant la migration et lorsqu’ils atteignent leurs destinations méridionales. La progéniture produite se développe pendant l’hiver chaud austral, avant de devenir adulte et de migrer vers le Nord (Trottier, 1971).
La migration nécessite non seulement des comportements spécifiques au stade adulte mais également des adaptations du développement des larves aquatiques. La prédation des poissons a une grande influence sur la survie et le développement des larves. La survie des larves sont plus élevée si les poissons insectivores sont absents. Les conditions climatiques peuvent favoriser la reproduction et la période de migration. (May et al., 2017)
Bibliographie
Beaton, G. (2007). Dragonflies & Damselflies of Georgia and the Southeast. University of Georgia Press, Athens, Georgia.
Corbet, P. (1999). Dragonflies Behavior and Ecology of Odonata. Ithaca, New York: Cornell University Press
Fraker, M., et B. Luttbeg. (2012). Predator-prey space use and the spatial distribution of predation events. Behaviour, 149(5)
Hopkins, G., Gall, B., et Brodie, E. (2011). Ontogenetic Shift in Efficacy of Antipredator Mechanisms in a Top Aquatic Predator, Anax junius (Odonata: Aeshnidae). Ethology, 117(12)
Needham, J.G. et Westfall, M.J. (1955) The Manual of the Dragonflies of North America (Anisoptera): Including the Greater Antilles and the Provinces of the Mexican Border. University of California Press, Berkeley and Los Angeles.
May, M.L., Gregoire, J.A., Gregoire, S.M., Lubertazzi, M.A., Matthews, J.H. (2017). Emergence phenology, uncertainty, and the evolution of migratory behavior in Anax junius (Odonata: Aeshnidae). PLoS ONE. 12(9)
Paulson, D. (2011). Dragonflies and Damselflies of the East. Princeton, New Jersey: Princeton University Press.
Pilon, J-G., et Lagacé, D. (1998). Les odonates du Québec. Chicoutimi, Canada: Entomofaune du Québec (EQ) Inc.
Robert, A. (1957). Les principales libellules du Québec (Vol. 7, N°9). Ottawa, Canada : Le Jeune Naturaliste.
Trottier, R. (1971). Effect of temperature of the life of Anax junius (Odonata : Aeshnidae) in Canada. The Canadian Entomologist. 103(12)
La similitude dans la largeur de la base et la longueur des ailes antérieures et postérieures fait d’Enallagma boreale un membre du sous-ordre Zygoptera (Resh, V.H., Cardé, R.T. (2009). Les caractéristiques principales qui définissent les Coenagrionidae sont le nombre de nervures transversales entre la base de l’aile et le nœud, dans l’espace Sc-R (moins de 5) (1), la marge occipitale dorso-postérieure de forme transverse (2), un CuA formé de deux cellules ou plus (3), l’absence de pseudostigmate (4) et l’absence de secteurs supplémentaires entre IR1 et RP2 (5); voir photos 2 et 3 (Garrison, R.W., von Ellenrieder, N., Louton, J.A. (2010)).
L’identification au genre a ensuite été possible en suivant la clé d’identification de Damselfly genera of the New World: an illustrated and annotated key to the Zygoptera (Garrison, R.W., et al (2010)) pour les femelles étant donné la présence d’un ovipositeur et d’une épine vulvaire. Parmi les caractéristiques qui ont permis de déterminer le genre (Enallagma) on retrouve: la cellule de forme quadrilatère à angle aigu (6), les bandes thoraciques parallèles au dos (7), le patron dorsal de la tête non tigré (8), l’abdomen pas majoritairement blanc (9), le segment S8 de l’abdomen possèdant une épine vulvaire (10) et l’ovipositeur ne dépassant pas le segment S10 (11); voir photos 2 à 4.
Pour identifier l’espèce (Enallagma boreale), deux ouvrages ont été utilisés soit Damselflies of the Northeast (Lam, E. (2004))et Dragonflies and Damselflies of the East (Paulson, D. (2011)) afin de comparer les différentes espèces du genre. Le choix s’est arrêté sur Enallagma boreale, car la distribution correspond mieux à la région où le spécimen a été trouvé, la taille des taches post-oculaires (12) est semblable et que l’ovipositeur est gros (11) chez le spécimen et chez l’espèce; voir figures 3 et 4.
Photo 2. Caractéristiques de l’aile permettant l’identification d’Enallagma boreale. Cliquer sur l’image pour une meilleure vue.
Photo 3. Caractéristiques de la tête et du thorax permettant l’identification d’Enallagma boreale. Cliquer sur l’image pour une meilleure vue.
Morphologie
Les adultes chez Enallagma boreale mesurent entre 28 et 36.6 mm et ont une envergure de 17 à 12 mm. Les femelles sont généralement plus grosses à cause des ovaires. L’espèce présente un dimorphisme sexuel ainsi qu’un polymorphisme chez la femelle. Le mâle étant principalement bleu et noir avec un patron de coloration se détaillant comme suit: une tête principalement bleue, un pronotum noir, un thorax bleu, des pattes bleues ou beige avec des bandes noires, un abdomen bleu avec trois bandes noires courtes et deux longues et un cerci noir. Les femelles présentent deux morphes: un semblable à celui du mâle et le second plutôt brun-roux. Le patron de marquage est semblable aux mâles mais avec un abdomen plus noir et des zones plus claires sur les trois derniers segments.3 La raison du polymorphisme n’a pas encore été découverte et l’hypothèse qui semblait la plus probable a été écartée. Celle-ci propose que les femelles bleues pourraient échapper aux mâles qui présentent un comportement agressif envers les femelles en période de reproduction. En effet, elle a été rejetée par une étude comparant le comportement du mâle envers les deux morphes de femelles qui s’est avéré être semblable peu importe leur couleur.4
Pour ce qui en est de la larve, elle présente un corps long et mince de couleur gris ou brun avec des branchies à l’extrémité postérieure de l’abdomen. Elle possède de grands yeux, un corps couvert de soie et un labium très développé permettant d’attraper ses proies.3 .
Distribution et habitat
Enallagma boreale est une espèce indigène de la zone néarctique et sa distribution s’étend d’est en ouest à travers le Canada et les États-Unis. Sa répartition nord-sud se définie au nord par la zone subarctique et au sud jusqu’à la Californie, l’Arizona et le Nouveau-Mexique 3. L’agrion boréal est l’espèce de zygoptère la plus nordique au Canada, vivant dans des températures estivales moyennes pouvant se situer aussi bas que 2-4°C. L’espèce peut se retrouver jusqu’à une altitude de 3500m (Corbet, P.S. (1999)).
La larve d’Enallagma boreale est aquatique et se retrouve dans les milieux lentiques d’eaux douces. À l’est, elle se retrouve dans les mares sans poisson, alors que dans l’ouest elle n’y est pas restreinte. Les adultes peuvent se trouver dans de multiples biomes terrestres (boisés, tourbières, marais, montagnes, etc.), mais restent à proximité des points d’eau.3 (Paulson, D. (2011)).
Reproduction et cycle de vie
Enallagma boreale est une espèce hémimétabole qui se reproduit pendant la saison estivale, particulièrement pendant les mois de juin et de juillet, mais la période de reproduction peut s’étendre de la fin du printemps jusqu’au début de l’automne dans les régions plus chaudes. Lorsque la femelle est prête, elle rejoint leur site de reproduction, en bordure des lacs ou d’étangs, où les males passent la plupart de leur temps.3 Suite à une parade nuptiale où le male exhibe ses couleurs et démontrent ses prouesses de vol, les deux individus adoptent une position d’accouplement particulière, mais caractéristique des zygoptères. Le male attrape la femelle par sa plaque mesostigmale à l’aide des appendices à l’extrémité de son abdomen. Il doit alors transférer son sperme jusqu’à ses glandes séminales, situées sous le second segment abdominal. La femelle va ensuite balancer son abdomen vers l’avant afin de rejoindre le second segment abdominal du mâle, où le transfert du sperme à lieu. La position qui en résulte, rappelant vaguement la forme d’un cœur, est appelée « roue de reproduction ». La copulation peut durer en moyenne 23 minutes.3
Par la suite, la femelle dépose ses œufs sur la végétation flottante, celle en bordure, ou même parfois directement dans l’eau. Le male aide la femelle lors du processus afin de s’assurer qu’aucun rival ne tente de féconder sa conquête. Brièvement après la ponte, le couple meurt. Les œufs peuvent éclore après seulement 11 jours à température élevée, mais peuvent prendre jusqu’à 61 jours à basse température. Les larves aquatiques sont minces et allongées, et donc bien adaptées aux courants rapides. Par contre, elles ne s’enfouissent pas dans la terre, contrairement aux larves de libellules. Ce stade peut durer quelques mois à quelques années, dépendamment des conditions, par exemple, les températures plus froides peuvent retarder le développement. De plus, les larves femelles nécessitent plus de temps pour se développer puisqu’elles une quantité de nourriture supplémentaire est nécessaire à la formation de leurs ovaires. Lors du passage entre le stade larvaire et le stade adulte, l’insecte sort de l’eau et s’accroche à la végétation, où la métamorphose peut prendre quelques jours. L’adulte vit en moyenne 4 jours, mais peut vivre jusqu’à 17 jours.5
Alimentation et prédation
Enallagma boreale est un redoutable prédateur qui s’alimente principalement de petits insectes au corps mou, comme les mouches et les moustiques. Les éphémères, les pucerons et certaines espèces de lépidoptères font également partie de leur régime. Les larves, elles aussi prédatrices, chassent de petits insectes aquatiques, tels que des larves d’éphémères, de diptères, ou encore des petits crustacés dulcicoles.3
L’agrion boréal est lui-même la proie de nombreux prédateurs. Notamment, les libellules représentent une menace constante, que ce soit pour les adultes ou pour les larves.6 Les adultes sont aussi victimes des araignées, des oiseaux, des Assilidés et même d’autres zygoptères. Les larves constituent la proie de grenouilles et d’insectes aquatiques, et doivent même redouter les autres larves d’Enallagma boreale, puisque les plus grosses font preuve de cannibalisme envers les plus petites et les œufs.7 En général, cette espèce n’a pas à craindre les poissons, puisqu’elle vit autour des points d’eau où ils sont absents. Par contre, si une population se retrouve dans une mare contenant des poissons, ces insectes représenteront une proie facile, puisqu’ils ne les considèrent pas comme des prédateurs potentiels.8
Corbet, P.S. (1999). Dragonflies: Behavior and Ecology of Odonata. Ithaca, États-Unis: Cornell University Press.
Garrison, R.W., von Ellenrieder, N., & Louton, J.A. (2010). Damselfly genera of the New World: an illustrated and annotated key to the Zygoptera. Baltimore, États-Unis: Johns Hopkins University Press.
Lam, E. (2004). Damselflies of the Northeast. Forest Hills, États-Unis: Biodiversity Books.
Paulson, D. (2011). Dragonflies and Damselflies of the East. Princeton, États-Unis: Princeton University Press.
Texte et photographies au domaine public CC0 1.0, sauf où précisé autrement
Figure 1. Vue d’ensemble de C. maculata
Spécimen capturé dans le chemin menant au lac Corriveau à la Station de biologie des Laurentides le 7 septembre 2017.
Classe: Insecta
Ordre: Odonata
Sous-ordre: Zygoptera
Famille: Calopterygidae
Genre: Calopteryx
Espèce: Calopteryx maculata (Beauvois, 1805)
Identification
Calopteryx maculata, de l’ordre Odonata, est une espèce du sous-ordre des zygoptères. En recherchant l’étymologie de ce nom, il est possible de trouver des racines grecque et latine. Calopteryx vient des mots grecs kalos et pteron, voulant respectivement dire belle et aile. Quant à maculata, ce terme tire son origine du mot latin macula signifiant « tache ». En effet, cela fait référence au ptérostigma blanc visible sur les femelles (BugGuide, 2014).
En premier lieu, ses yeux disproportionnés par rapport à la tête sont éloignés l’un de l’autre (University of Florida, 2015). En considérant également les petites antennes, ces diverses caractéristiques permettent de placer le spécimen au sein de l’ordre des odonates (voir figure 2). À première vue, il est possible de remarquer les ptérostigmas colorés en blanc placés en contraste sur chacune des deux paires d’ailes brunes (voir figure 4). Cette caractéristique permet d’identifier notre spécimen comme étant une femelle, puisque les mâles de cette espèce n’ont pas de ptérostigmas. De plus, l’abdomen de couleur noir du spécimen (voir figure 1) confirme le sexe de l’individu puisque le mâle arbore plutôt un abdomen bleu ou vert métallique (BugGuide, 2014).
Figure 2. Vue complète de la tête de C. maculata
Deuxièmement, il importe de statuer le sous-ordre auquel appartient ce spécimen. Cette partie de l’identification est simplifiée par la division dichotomique de l’ordre des odonates en deux sous-ordres: Zygoptera et Anisoptera. Malgré le fait d’avoir deux paires d’ailes de taille quasi identiques, zygoptera se différencie de son groupe frère par le point d’attache des ailes au thorax qui est plus étroit (voir figure 3). Par ailleurs, les yeux fortement décollés, tel que mentionné ci-haut, confirment l’appartenance du spécimen aux zygoptères.
Figure 3. Base de l’aile étroite (Zygoptera) et non pétiolée (Calopterygidae) de C. maculata
Troisièmement, il faut identifier la famille à laquelle le spécimen appartient. La base des ailes constitue la caractéristique clé. En effet, celle de notre demoiselle n’est pas pétiolée (voir figure 3), c’est-à-dire que la base de l’aile n’est pas rétrécie. Cette observation témoigne de son appartenance à la famille des Calopterygidae, puisqu’il s’agit de la seule famille de demoiselles dont la base des ailes est non pétiolée (Beckemeyer et Huggins, 1998). Cependant, un facteur plus quantitatif renforce cette identification à cette famille. La technique consiste à compter le nombre de nervures transversales entre la base de l’aile et le nœud (University of Florida, 2015). Tout d’abord, le nœud est représenté par la partie supérieure de l’aile qui s’enfonce légèrement. La seule famille à avoir environ dix nervures transversales ou plus est celle des Calopterygidae. Cette situation s’applique à notre spécimen qui en a approximativement 23 (voir figure 4). De ce fait, les autres familles de zygoptères ne répondent plus aux critères d’identification. Ces dernières ne possèdent environ que deux de ces nervures transversales.
Figure 4. Vue d’ensemble de l’aile du spécimen (femelle)
Finalement, il est important de noter que la demoiselle en question n’a pas de taches rouges à la base de ses ailes. Cela contribue à définir le genre, car cette caractéristique est propre au genre Hetaerina. Cette hypothèse écartée, la demoiselle ne peut donc qu’appartenir au genre Calopteryx. Pour définir l’espèce à laquelle elle fait partie, les ailes foncées allant du brun au noir est un indice majeur. Il faut noter que parmi toutes les espèces du genre Calopteryx, la seule à avoir cette particularité est Calopteryx maculata.
Basé sur toutes ces caractéristiques morphologiques et anatomiques, il est effectivement possible de conclure que ce spécimen appartient à l’espèce C. maculata. Le caractère le plus marquant est le nombre de nervures transversales entre la base de l’aile et le nœud qui permet d’identifier à la famille Calopterygidae. Aussi, les ailes brunes sont assez uniques parmi le genre Calopteryx. De plus, l’identification à l’espèce pour l’ordre des odonates se fait majoritairement grâce aux caractéristiques clés se retrouvant sur les ailes.
Répartition géographique/ Habitat
La demoiselle bistrée (Dubuc, 2007, p. 43), comme elle est communément appelée, peut mesurer de 39 à 57 mm. Elle se trouve principalement dans les régions néarctiques, plus spécifiquement dans le Sud du Manitoba, du Sud jusqu’à l’Est du Texas, de l’Est jusqu’à la Nouvelle-Écosse et la Floride (Evans, 2007, p.69). Pour ce qui est de son habitat, celle-ci se retrouve perchée sur des branches situées aux alentours de ruisseaux ou de rivières lors des journées ensoleillées. En cas de journée plus froide, elles peuvent être présentes dans les forêts entourant les cours d’eau, comme le témoigne le spécimen. La larve, contrairement à l’adulte, est complètement aquatique. Cette dernière se niche près des plantes aquatiques. Il s’agit d’une espèce qui est principalement présente durant la période de mai jusqu’à juillet, voire août (BugGuide, 2014).
Cycle de vie
Étant une espèce diurne, C. maculata s’accouple habituellement en début d’après-midi. Lors de la reproduction, les mâles se livrent à une féroce compétition pour l’accès aux femelles. Ceux-ci adoptent un comportement territorial pour offrir aux femelles le meilleur lieu de fécondation suite à l’accouplement. Tout d’abord, le mâle a un organe de reproduction secondaire situé en dessous de son thorax. Avant l’accouplement, il transfère sa semence de son ouverture génitale à cet organe. De plus, il est possible de confirmer que notre spécimen est effectivement une femelle par l’absence des ces pinces anales au bout de son abdomen (voir figure 5). Pour attirer la femelle, le mâle se positionne en croix, les ailes postérieures en angle droit avec le corps. Cette technique permet de relever l’abdomen pour attraper celle-ci par son cou. Grâce à ses pinces anales, le mâle peut ainsi maintenir la femelle en place. La femelle, quant à elle, ramène son abdomen vers l’avant pour joindre le bout à l’organe reproducteur secondaire du mâle. Cette position forme ainsi une structure semblable à une roue (voir figure 6) (Evans, 2007, p.60-61).
Figure 5. Le bout de l’abdomen du spécimen (femelle) sans pinces anales
Aussitôt fécondée, la mère se dépêche de pondre. Les œufs de Calopteryx maculata se retrouvent soit flottants dans l’eau, émergés ou submergés par de la végétation. La dernière option est favorisée par ces demoiselles, puisqu’il s’agit d’un milieu optimal pour protéger les progénitures. Par ailleurs, le mâle vole à proximité pour tenir la garde contre d’éventuels prédateurs. Lorsqu’un mâle s’accouple avec plusieurs femelles, un agrégat d’œufs se forme en un même lieu, car celui-ci doit leur offrir une protection. Enfin, la larve, nommée naïade, sort de sa coquille 10 à 120 minutes plus tard (NatureServe Explorer, 2016).
Figure 6. Positions du mâle (droite) et de la femelle (gauche) lors de l’accouplement de C. maculata Kevin Payravi, CC BY-SA 3.0, Wikimedia Commons
Cette espèce témoigne du même cycle de vie que tous les autres odonates, c’est-à-dire d’un développement hémimétabole, définit par la métamorphose incomplète. Alors que les larves se dédient à la croissance, les adultes se reproduisent. La durée de ce cycle de vie est d’environ un à deux ans, dont la majorité a lieu lors du stade larvaire. Fait intéressant, les demoiselles respirent par l’anus. En effet, Evans (2007, p.60-61) explique que les larves ont trois feuillets de branchies situés sur le bout de l’abdomen qui permettent leur respiration. Afin d’augmenter leur taille, les demoiselles en devenir font la mue de dix à douze fois, où elles acquièrent une pigmentation plus foncée. À la dernière métamorphose, l’adulte juvénile émerge et rejoint la terre ferme. On lui accorde le terme « adulte juvénile », puisque sortant de l’eau, il n’a pas encore atteint sa maturité sexuelle (Evans, 2007, p.60-61). À cette étape, le nouvel individu finit de durcir ses ailes et son abdomen avant de pouvoir s’envoler. Par la suite, la demoiselle adulte vit de quelques semaines à quelques mois le temps de se reproduire (North American Insects & Spiders).
C. maculata est une espèce totalement carnivore et prédatrice. En effet, elle se nourrit de petits insectes, tant au stade larvaire qu’au stade adulte (BugGuide, 2014). Cependant, elle ne se trouve pas à la tête de la chaîne alimentaire. Comme la presqu’entièreté des insectes, elle est la proie des autres animaux. Certains facteurs influencent la durée de vie de l’adulte. Comme l’indique Stoks & Cordoba-Aguilar (2012), le risque de mortalité croît en présence de parasites, de prédateurs, de mauvaises conditions environnementales et du manque de nourriture. Aussi, le harcèlement des mâles pour s’accoupler est un facteur dangereux pour les femelles.
Bibliographie
Beckemeyer, RJ., Huggins, DG (1998). The Kansas School Naturalist, Emporia State University, vol. 44
Ce spécimen peut être classé dans l’ordre des Odonata de par le fait qu’il possède des ailes non réduites et qui ne peuvent pas se replier sur le corps. Puisqu’elles sont perpendiculaires au corps au repos et que les postérieures sont plus larges à la base que les antérieures, l’insecte est un anisoptère.
La famille des Aeshnidae est caractérisée par trois éléments morphologiques (2):
-Les triangles formés par les nervures des ailes antérieures et postérieures sont placés à égale distance de l’arculus, soit la séparation de la troisième nervure en plusieurs nervules. De plus, les triangles sont orientés vers l’apex de l’aile de la même façon;
-Le lobe médian du labium est entier;
-Les yeux se touchent sur la ligne médiodorsale.
Les membres du genre Aeshna possèdent un thorax noir ou brun, avec des taches brunes, jaunes ou vertes. Leurs segments abdominaux n’ont qu’une seule carène pour chaque côté. Les Aeshna ont tous une caractéristique particulière au niveau de l’aile: la médiane antérieure et la branche asymétrique qui y est associée forme une fourche qui se dirige vers le bord antérieur de l’aile diagonalement (2).
En général, les membres d’Aeshna sont parmi les plus grands anisoptères vivants. Leur grande taille, ainsi que la coloration particulière de leur corps, sont des caractères suffisants pour une identification préliminaire.
Aeshna umbrosa mesure en moyenne 7,5cm en longueur (1) et leurs ailes mesurent plus que 40mm en longueur. Les membres de l’espèce ont une bande frontoclypéale (soit entre le front et le clypéus, un sclérite qui délimite la marge inférieure de la face) mince et brunâtre, voire de la même couleur que le front et le clypéus.
Cette espèce se distingue des autres Aeshna avec ses deux bandes latérales situées sur le ptérothorax et aucune tache jaune entre les deux bandes. Ces deux bandes latérales sont rectilignes, étroites et de couleur jaune, verte ou bleue, selon le sexe, entourées de noir. Les mâles possèdent des yeux turquoise, ainsi que des bandes dont la couleur varie du bleu-vert au jaune-vert. Les taches abdominales sont très petites et sont de couleur bleue ou verte. Les femelles quant à elles ont des yeux bruns avec des bandes latérales jaune-verdâtre ou des yeux bleutés avec des bandes vert-jaunâtre. Les femelles ont des taches abdominales strictement bleues (3) (4).
Aeshna tuberculifera ressemble beaucoup à Aeshna umbrosa, mais ses bandes ne sont pas entourées de noir et sont plus larges. Le 10e segment abdominal d’A. umbrosa possède des taches ternes, en contraste au 10e segment entièrement noir d’A. tuberculifera. De plus, ce segment possède une saillie épineuse chez A. tuberculifera, alors qu’A. umbrosa en est dépourvu.
On retrouve deux sous-espèces d’Aeshna umbrosa: Aeshna umbrosa umbrosa et Aeshna umbrosa occidentalis. La première possède des taches abdominales vertes plus petites sur les segments postérieures, tandis que la deuxième possède des taches bleues et plus grandes (3).
Habitat
Comme son nom l’indique, l’aeschne des pénombres peut être trouvée dans des endroits avec un peu d’ombre et où l’eau ne bouge pas ou très peu. Cela inclut les lacs, les étangs et les marécages. Cependant, il n’est pas rare de la trouver dans une zone dégagée, surtout lorsqu’elle chasse. C’est dans un tel lieu que nous avons capturé notre spécimen. Pour ce qui est des larves, elles peuvent être trouvées dans les cours d’eau mentionnés précédemment (5).
Diète et techniques de chasse
Toutes les espèces d’odonates sont, sans aucune exception, de redoutables carnivores et ce, au stade larvaire et adulte. Les larves mangent en général les larves des insectes aquatiques, mais peuvent aussi se nourrir de d’autres animaux aquatiques, tels que des crustacés d’eau douce, des tétards et même, des poissons de petite taille. Les adultes quant à eux, se nourrissent de tout insecte ayant une plus petite taille que l’odonate et cela inclut les moustiques, mouches, papillons, coléoptères et même autres odonates. Une particularité de la diète des odonates est qu’ils ne consomment pas les ailes; ils arrachent les ailes de leurs proies avant de les dévorer. On peut dire d’eux qu’ils sont des prédateurs opportunistes (5) (6).
Aeshna umbrosa chasse le plus souvent aux alentours du coucher du soleil, profitant de sa coloration pour se camoufler. En général, les odonates possèdent deux principales techniques de chasse: hawking et gleaming. La première technique consiste en une capture dans les airs d’un autre insecte en vol. La capture se fait principalement avec la bouche, mais peut aussi être complimentée des pattes thoraciques pour bien maintenir la proie. Avec la technique de hawking, Aeshna umbrosa peut consommer quotidiennement jusqu’à 20% de sa masse corporelle (5). La deuxième technique consiste en un survol des zones avec beaucoup de végétation. Lorsque l’odonate aperçoit une proie potentielle, elle plonge vers la proie et l’attrape avec ses pattes thoraciques. Une autre technique peut être utilisée par certaines espèces d’odonates: un individu peut se percher sur un endroit quelconque et attend patiemment le passage d’une proie avant de plonger sur cette dernière (6).
Un odonate en chasse effectue beaucoup de mouvements de va-et-vient afin de patrouiller les environs. Il n’est pas rare d’apercevoir une nuée d’odonates de la même espèce, ou de différentes espèces, qui chassent ensemble. Cette agglomération est due à la présence d’un autre essaim important de potentielles proies dans les parages. Aeshna umbrosa démontre cette caractéristique de former un essaim pour se nourrir (5) (6).
Développement larvaire
Pour réussir à passer à travers l’hiver, les œufs des odonates tombent en diapause, un ralentissement métabolique qui assure leur survie jusqu’au printemps. C’est à ce moment qu’ils commencent à éclore pour libérer des larves (aussi appelées naïades) qui finissent par mesurées entre 3,8 et 4,4 centimètres chez Aeshna umbrosa. Après une période de croissance dans l’eau, elles émergent et passent d’une respiration par branchies anales à une respiration par spiracles thoraciques. Pour compléter leur dernière mue, les larves avalent de l’eau qui les fait gonfler, la cuticule cédant sous la pression qui s’accumule. Les ailes nécessitant une période de sclérification avant de permettre le vol, les odonates sont particulièrement vulnérables à cette période du développement. La mue vers le stade adulte se produit généralement de nuit afin de minimiser la prédation (7).
Distribution
Aeshna umbrosa est répartie à travers le continent nord-américain, dans le sud de la région boréale, de la côte Pacifique jusqu’à la côte Atlantique excluant la province de Terre-Neuve. Cette aeschne se trouve aussi aux États-Unis, allant du Canada jusqu’à la région sud-américaine exclusivement, où le climat sec empêche toute dispersion de l’espèce. La sous-espèce umbrosa est distribuée à l’est du continent, tandis que la sous-espèce occidentalis se trouve à l’ouest (3) (4).
Références
1. « Species Aeshna umbrosa » (en ligne), Iowa State University. 2003-2014. Page consultée le 20 octobre 2014 sur BugGuide.net.
2. Pilon Jean-Guy et Lagacé Denise, 1998. Les odonates du Québec. Entomofaune du Québec (EQ) Inc., 367 pages. p.31, 38-39, 59-64.
3. Paulson Dennis , 2011. Dragonflies and Damselflies of the East. Princeton University Press, 538 pages. p.199-200.
4. Paulson Dennis, 2009. Dragonflies and Damselflies of the West. Princeton University Press, 536 pages. p.222-223.
5. Dunkle Sidney W., 2000. Dragonflies Through Binoculars. Oxford University Press, 369 pages.
Lieu de capture : dans les herbes d’un fossé de drainage, en bordure d’une route en terre battue, à proximité d’une tourbière boisée de la station de biologie des Laurentides. Outil de capture : filet fauchoir Date de capture : 1 septembre 2016 Préparation: d’abord placé dans un récipient de chasse contenant un mélange d’acétate d’éthyle et de plâtre pour le tuer rapidement, avant d’être mis à sécher dans une enveloppe de plastique avec carton
Identification, classification et morphologie
Figure 1 : Front blanc du mâle Sympetrum obtrusum capturé. Photo réalisée par Étienne R.-Dionne.
Frappante, la couleur blanche du front des mâles est l’une des caractéristiques principales permettant de distinguer S. obtrusum parmi les espèces du genre Sympetrum, d’où leur nom vernaculaire “White-faced Meadowhawk”.
Généralement, les individus de cette espèce mesurent de 30 à 35mm. Ils ont les ailes claires. Mâles et femelles présentent un certain dimorphisme. En effet, les femelles n’ont pas le front blanc caractéristique des mâles ni l’abdomen teinté de rouge. Celles-ci, comme les juvéniles, ont plutôt la face d’une couleur qui tire sur le jaune, le brun ou le vert, ainsi qu’un abdomen cendré, ce qui les rend difficiles à distinguer de S. rubicundulum et S. internum (Dunkle, 2000).
Figure 2 : Pièces génitales. A : Appendices anaux supérieurs de l’extrémité de l’abdomen mâle – B : Échancrure de l’hameçon du deuxième segment de l’abdomen mâle – C : Huitième sternite horizontale de l’extrémité de l’abdomen femelle (Ovipositeur) – D: Lames vulvaires du neuvième segment de l’abdomen femelle. Croquis réalisé par Miguelly Bélanger. Inspiré de Robert (1963).
Comme les S. obtrusums, le spécimen capturé possède une face blanche. Cependant, ce sont les appendices anaux supérieurs de l’extrémité de l’abdomen ainsi que l’échancrure de l’hameçon du 2e segment de l’abdomen qui a permis de confirmer son identification (Robert, 1963).
De plus, la position médiane du nodus ainsi que la disposition des nervures principales du supertriangle de l’aile postérieure de notre spécimen sont typiques des Sympetrum (Pilon & Lagacé, 1998). Appartenant également à la famille des libellulidés, leurs ailes postérieures présentent un champ anal en forme de pied possédant un « orteil » plutôt développé. Qui plus est, l’asymétrie de la base des ailes antérieures et postérieures (la base des ailes antérieures étant plus large que celle des ailes postérieures) corrobore l’appartenance au sous-ordre des anisoptères (Silsby, 2001).
Figure 3 : Aile postérieure caractéristique du genre Sympetrum représentant le nodus (A), le pterostigmate (B), le supertriangle et le champ anal. Croquis réalisé par Miguelly Bélanger. Inspiré de Robert (1963).
Dans une perspective d’ensemble, les libellules présentent plusieurs caractéristiques distinctives (Pilon & Lagacé, 1998). Elles ont de courtes antennes. Leurs yeux composés sont de si grandes tailles par rapport au volume de leur tête, qu’ils se rejoignent au sommet de la capsule céphalique. Ils possèdent un long et mince abdomen. Leur thorax orienté vers l’avant permet une capture de proies efficace, en les emprisonnant dans leurs pattes. Au repos, les ailes se posent à plat. Leurs larves possèdent un masque labial étonnant, leur permettant d’atteindre leurs proies avec grande vélocité (Kalkman & al., 2000). Voici un ensemble de caractères partagés par l’espèce du spécimen capturé.
Figure 4 : Taxonomie hiérarchique de Sympetrum obtrusum. Inspiré de Dunkle (2000) et Cresswell & Block (2016).
Les libellules à travers les âges et les continents
Répartis sur l’ensemble de la planète depuis des millénaires, les odonates ont intéressé les hommes et suscité une fascination culturelle à travers les âges (Pilon, 1998). Venant du grec ancien, le mot « odonata » fut créé par Fabricius (1793) pour désigner les « mandibules munies de dents », alors que le mot « Anisoptera » souligne l’asymétrie des ailes (Pilon & Lagacé, 1998). Ceci dit, c’est Linné (1758) qui a introduit le terme « libellules ». En extrême Orient, les libellules sont plutôt bien considérées. Elles y ont d’ailleurs longtemps été utilisées pour traiter les inflammations de la gorge. Au Japon, elles symbolisent la force, la bravoure et la victoire. En Europe, au contraire, ces insectes sont considérés comme maléfiques, une croyance populaire probablement tirée du moyen âge. Pour cette raison, ils se faisaient même appeler « tire-z’yeux » ou « crève-œil » (d’Aguilar, 1985). Plus près d’ici, on a souvent considéré à tort les libellules comme des insectes piqueurs. Aujourd’hui encore, les libellules suscitent un certain engouement.
Évolution
Figure 4 : Représentation des Ères géologiques. Inspirée de Cohen & al. (2013).
Apparus durant la période du carbonifère, les Protodonata sont peut-être les ancêtres des Odonates. Datant du permien, les libellules sont parmi les plus anciens insectes volants (paléoptère) (Grimaldi, D. et Engel, 2005). Dans le même taxon, Protodonata, se trouve le plus grand insecte connu à ce jour (Méganeuropsis permiana), d’une envergure d’ailes de 70cm (Beckemeyer, 2000). Des fossiles de zygoptères et d’anisoptères (Odonata au sens strict) ont d’abord été trouvés au Mésosoïque: les premiers fossiles d’anisoptères modernes datent du Trias (Silsby, 2001). Ceux-ci auraient donc déjà été présents avant que la Pangée se soit complètement disloquée! Émergeant de cette lignée, les libellulidés sont apparus lors de l’Éocène. Avec leurs ailes puissantes et leur corps effilé, leurs ancêtres ont pu bénéficier d’un avantage évolutif important, leur permettant de se distribuer sur un large territoire. À ce jour, on compte près de 7000 espèces d’Odonates (Kalmman & al., 2008) et plus de 60 espèces de Sympetrum à travers le monde (Pilgrim & Dohlen, 2007).
Répartition et habitat
Figure 5 : Carte de la distribution de l’espèce Sympetrum obtrusum. Carte réalisée par Miguelly Bélanger. Inspirée de Dunkle (2000).
Espèce indigène de l’Amérique du Nord, Sympetrum obtrusum se retrouve dans la région zoogéographique Néarctique. Il est principalement recensé au sud du Canada et au nord des États-Unis, mais semble absent du Yukon et du sud de l’Amérique du Nord (Dunkle, 2000; Catling, 2003). Dans ces territoires, il occupe une variété de milieux humides (étangs, lacs, cours d’eau lentiques) et spécialement les tourbières et les marais (Pilon et Lagacé, 1998). Considérant la dispersion des adultes à partir des étendues d’eau, certains avancent que les individus matures constituent une population distincte de celle des larves, malgré le fait que l’un et l’autre occupent les mêmes zones (Faydenko, 2013).
Cycle de vie et alimentation
Essentiels au développement des libellules, les points d’eau douce permettent le développement de leurs œufs et de leurs larves (Kalkman et al., 2008). Généralement, les œufs éclosent environ trois semaines après la ponte, sauf ceux pondus à la fin de la saison estivale, qui tombent en dormance durant l’hiver (Silsby, 2001). C’est d’ailleurs sous forme d’œufs que les embryons de Sympetrum obtrusum passent l’hiver, pour voir leurs larves éclore au printemps et les adultes émerger fin juin, début juillet (Walker & Corbet, 1975). Ainsi, deux générations se succéderont chaque année. Après éclosions, plusieurs stades larvaires s’enchaineront, entrecoupées de mues. Chez les libellules, le premier stade (stade protolarvaire) est caractérisé par l’absence de patte et ne dure qu’une courte période de temps (de quelques secondes à quelques heures) (Silsby, 2001). Par suite, certaines larves de libellules seront plus passives et d’autres plus actives dans leur comportement de chasse, mais toutes sont prédatrices. Grâce à leur masque labial, qui leur permet d’attraper leurs proies à grande vélocité, les larves des Odonates arrivent à se saisir d’insectes, de têtards et même de petits poissons (Corbet, 1999).
Video 1 : Larve de libellule se nourrissant d’une larve de coléoptère à l’aide de son masque labial (par Derek Wheaton, 2015). CC BY-SA 4.0
Contrairement à certains autres genres, le genre Sympetrum consomme non seulement des proies à chacun de ses stades larvaires, mais passe aussi la majeure partie de son cycle de vie sous l’eau (Pritchard, 1964; Corbet, 1999). Les larves de libellules prennent de quelques semaines à quelques années pour se développer, selon l’espèce et différents facteurs environnementaux, tel que la température de l’eau et la quantité de nourriture disponible (Corbet, 1999; Silsby, 2001). Au dernier stade larvaire avant la métamorphose, les derniers organes vitaux finissent de se développer et la larve se prépare pour une dernière mue. L’émergence des libellules matures se fait à l’extérieur de l’eau, sur la végétation ou sur la rive, au courant de la nuit ou au matin, lorsque l’air a pu se réchauffer (Silsby, 2001).
Video 2 : Émergence d’une libellule après métamorphose (auteur inconnu, 2010). CC BY-SA 4.0
Après avoir quitté leur site d’émergence, les libellules occupent la majeure partie de leur temps à chasser, principalement des diptères et de petits insectes volants (Pilon & Lagacé, 1998; Dunkle, 2000). Avec les zygoptères, les libellulidés se distinguent des autres Odonates de par leurLe sympétrum éclaireur démontre une (édité 2018-01-22) mode de chasse à l’affût, repérant leurs proies à partir d’un perchoir, avant de se jeter sur elles (Pilon & Lagacé, 1998). En une seule journée, une libellule peut ainsi consommer jusqu’au cinquième de son poids (Powell & Twist, 1999). Selon les conditions, les libellules en développement auront besoin d’une semaine à un mois pour s’alimenter. Une période cruciale où elles se disperseront afin d’acquérir l’énergie suffisante au développement de leurs organes sexuels (Aguesse,1968; Corbet, 1952). À la recherche de ressources, les mâles Sympetrum obtrusum sont très abondants dans les prairies de Carex. Lors de cette phase de développement, il est possible d’y en observer plus d’un par 10 pi2 (Paulson, 2011) ! Ce n’est qu’en tant qu’individu mature que les mâles et femelles retourneront près de points d’eau pour s’accoupler (Corbet, 1999).
Reproduction
Chez les libellules, le mécanisme de transfert de sperme est indirect (Kalkman, 2008). Après avoir été produit dans les testicules, situés au bout de l’abdomen, le sperme est transféré de manière externe à l’organe copulatoire mâle, à la base de l’abdomen. Grâce à un hameçon, le mâle retient l’abdomen de la femelle durant la copulation. Ainsi, il insémine la femelle et en profite même pour retirer le sperme de ses compétiteurs. Cette compétition spermatique chez les Odonates a été reportée pour la première fois par Waage (1979). Aujourd’hui, c’est ce qui en fait l’un des groupes animaux les plus étudiés pour leur comportement reproducteur (Kalkman, 2008). Suite à la copulation, le mâle reste accroché à la femelle jusqu’à la ponte (Silsby, 2001). Les perforations sur l’une des extrémités des œufs oblongs permettent aux spermatozoïdes de les pénétrer. Puis, les œufs sont ensuite rapidement relâchés à la surface de l’eau ou sur le sol environnant, la femelle ayant un ovipositeur vestigial.
Utilité pour l’homme et statut de conservation
Très sensibles à la qualité de leur habitat et aux perturbations environnementales, les libellules sont souvent utilisées pour évaluer l’ampleur des changements environnementaux, sur le court et le long terme. Aussi, elles sont utilisées comme bioindicateurs de la santé de l’écosystème, dans les projets de conservation et de restauration (Kalman, 2008; Clark & Samways, 1996). Le réchauffement climatique, l’agriculture, l’introduction d’espèce exotique, l’urbanisation et plus encore, entraînent cependant la dégradation et la perte de diversité des milieux humides, dont dépendent les Odonates (Hassall & Thompson, 2008; Kadoya & al., 2009; Rodriguez & al., 2005). De fait, certaines espèces de libellules sont aujourd’hui dans une situation critique (Suh & Samways, 2005; Hämäläinen, 2004). Considérant la large gamme d’organismes interagissant avec les libellules, que ce soit au stade larvaire ou adulte, il est d’autant plus important de protéger les milieux humides, dont ils dépendent.
Bibliographie
Beckemeyer RJ. 2000. The Permian insect fossils of Elmo, Kansas. Emporia State University.
Bulankova E. 1997. Dragonflies (Odonata) as bioindicators of environment quality. Biologia, Bratislava 52:177-180.
Catling PM. 2003. Dragonflies (Odonata) of the Northwest Territories: Status Ranking and Preliminary Atlas. Northwest Territories, Environment and Natural Resources.
Clark TE, Samways MJ. 1996. Dragonflies (Odonata) as indicators of biotope quality in the Kruger National Park, South Africa. Journal of applied ecology:1001-1012.
Cohen K, Finney S, Gibbard P, Fan J-X. 2013. The ICS international chronostratigraphic chart. Episodes 36:199-204.
Corbet PS. 1952. An adult population study of Pyrrhosoma nymphula (Sulzer):(Odonata: Coenagrionidae). The Journal of Animal Ecology:206-222.
Corbet. 1999. Dragonflies: behaviour and ecology of Odonata. Harley Books.
Cresswell S, Block N. 2016. Species Sympetrum – White-faced Meadowhawk. (2016; http://bugguide.net/node/view/8056)
d’Aguilar J, Dommanget J-L. 1985. Guide to the dragonflies of Europe and North Africa. Delachaux et Niestlé.
Dunkle SW. 2000. Dragonflies through binoculars: a field guide to dragonflies of North America. Oxford University Press.
Fabricius JC. 1793. Entomologia systematica emendata et aucta.
Faydenko JL, Butler LM, Swanson BJ. 2013. As the Dragon Flies: Population Structure in Sympetrum obtrusum Dragonflies.
Grimaldi D, Engel MS. 2005. Evolution of the Insects. Cambridge University Press.
Hagen H. 1867. Revision der von Herrn Uhler beschriebenen Odonaten. Stett. Ent. Ztg. 28: 87–95. Sympetrum obtrusum.
Hämäläinen M. 2004. Critical species of Odonata in the Philippines. International Journal of Odonatology 7:305-310.
Hassall C, Thompson DJ. 2008. The effects of environmental warming on Odonata: a review. International Journal of Odonatology 11:131-153.
Kadoya T, Suda S-i, Washitani I. 2009. Dragonfly crisis in Japan: a likely consequence of recent agricultural habitat degradation. Biological Conservation 142:1899-1905.
Kalkman VJ, Clausnitzer V, Dijkstra K-DB, Orr AG, Paulson DR, van Tol J. 2008. Global diversity of dragonflies (Odonata) in freshwater. Hydrobiologia 595:351-363.
Labhart T, Nilsson D-E. 1995. The dorsal eye of the dragonfly Sympetrum: specializations for prey detection against the blue sky. Journal of Comparative Physiology A 176:437-453.
Paulson D. 2011. Dragonflies and Damselflies of the East. Princeton University Press.
Perron J, Jobin L, Mochon A. 2005. Odonatofaune du parc national de la Yamaska, division de recensement de Shefford, Québec. Le Naturaliste canadien 129:17-25.
Pilgrim EM, Von Dohlen CD. 2007. Molecular and morphological study of species-level questions within the dragonfly genus Sympetrum (Odonata: Libellulidae). Annals of the Entomological Society of America 100:688-702.
Pilon J-G, Lagacé D. 1998. Les odonates du Québec: traité faunistique. Entomofaune du Québec.
Powell D, Twist C. 1999. A guide to the dragonflies of Great Britain. Arlequin Press.
Pritchard G. 1964. The prey of dragonfly larvae (Odonata; Anisoptera) in ponds in northern Alberta. Canadian Journal of Zoology 42:785-800.
Robert A. 1963. Les libellules du Québec. Québec: Station biologique du Mont Tremblant, Service de la recherche, Ministère de la chasse et des pêcheries, 1963, 1966 printing.
Rodríguez C, Bécares E, Fernández-Aláez M, Fernández-Aláez C. 2005. Loss of diversity and degradation of wetlands as a result of introducing exotic crayfish. Biological Invasions 7:75-85.
Silsby J. 2001. Dragonflies of the World. Csiro publishing.
Suh AN, Samways MJ. 2005. Significance of temporal changes when designing a reservoir for conservation of dragonfly diversity. Biodiversity & Conservation 14:165-178.
Waage JK. 1979. Dual function of the damselfly penis: sperm removal and transfer. Science 203:916-918.
Walker E, Corbet P. 1975. The Odonata of Canada and Alaska. Vol. 3, Part III: The Anisoptera—Three Families: Toronto: University of Toronto Press.
Figure 1 : Photographie d’un Sympetrum vicinum capturé à la Station biologique des Laurentides le jeudi 03 septembre 2015
I- Identification du spécimen et morphologie
L’identification de notre spécimen a été possible grâce à la clé d’identification des libellules du Québec (Adrien Robert, 1963).
Tout d’abord, il faut distinguer deux sous-ordres dans l’ordre des Odonates : les anisoptères et les zygoptères. On peut les reconnaître grâce à la position et à la taille de leurs ailes. En effet, les zygoptères possèdent des ailes antérieures et postérieures de forme et de taille semblables qui, au repos, sont repliées l’une contre l’autre. Les anisoptères, quant à eux, ont des ailes postérieures plus larges à la base que leurs ailes antérieures. La figure 1 montre bien qu’il s’agit d’un insecte du sous-ordre des anisoptères.
Le triangle des ailes antérieures et le triangle des ailes postérieures n’ont pas la même orientation spatiale. De plus, la distance entre l’arculus et le triangle des ailes antérieures est plus grande que celle des ailes postérieures. Les nervures antodénales forment également des lignes continues, symétriques par rapport à la nervure sous-costale.
Figure 2 : Photographie des ailes antérieures et postérieures de Sympetrum vicinum
Le triangle des ailes antérieures à une forme étroite. L’angle anal des ailes postérieures est arrondi chez le mâle comme chez la femelle et il n’y a pas d’auricule. La courbure anale de l’aile postérieure est de la forme d’une botte. Les ailes sont transparentes sans reflet métallique. Il s’agit donc de la famille des Libellulidés.
Figure 3 : Photographie des ailes de Sympetrum vicinum afin de montrer où se trouvent la courbure anale et l’angle anal
La branche de la médiane M2 n’a pas de sinuosité vers son milieu et sur les ailes postérieures la première nervure cubitale (Cu1) commence à l’angle caudal du triangle. De plus les ailes sont colorées en brun à leur base et sont entièrement hyalines.
Figure 4 : Emplacement des nervures M2 et Cu1 sur l’aile postérieure de Sympetrum vicinum
On peut aussi remarquer que la face de notre spécimen est brune et jaune. Il s’agit donc du genre Sympetrum.
Figure 5 : Photographie de la tête de Sympetrum vicinum
Les côtés du thorax de la libellule sont rouges et le bord ventral des appendices anaux supérieurs n’ont pas de dents. Il s’agit donc de Sympetrum vicinum.
Figure 6 : Photographie du thorax de Sympetrum vicinum
Figure 7 : Photographie des appendices anneaux de Sympetrum vicinum
II- Classification
Notre spécimen appartient à l’espèce Sympetrum vicinum selon la classification suivante:
Règne: Animalia
Embranchement: Arthropoda
Sous-embranchement: Hexapoda
Classe: Insecta
Ordre: Odonata
Sous-ordre: Anisoptera
Famille: Libellulidae
Genre:Sympetrum
Espèce: Sympetrum vicinum (Hagen, 1861)
III- Habitat
Sympetrum vicinum est l’une des espèces d’anisoptères les plus nombreuses en Amérique du Nord. Au Québec, elle est principalement trouvée au sud où se situent les érablières à caryer cordiforme et à tilleul. Les basses terres du Saint-Laurent en sont donc très peuplées (Savard, 2013).
Elle favorise comme habitat des emplacements où la nourriture est abondante pour les larves. Selon une étude de l’odonatofaune du parc national de la Yamaska effectuée en 2005, les Sympetrum vicinum étaient présentes autour du réservoir Choinière, du ruisseau Fleurant-Messier, de l’étang à castor, des mares (peu ou très eutrophiées) et des champs en friche. En revanche, elles étaient absentes de la rivière Yamaska Nord, témoignant de la nécessité d’un courant faible ou absent pour leur développement (Perron et al, 2005). Cette espèce est donc qualifiée d’ubiquiste puisqu’elle peut habiter plusieurs types d’habitat autour de plans d’eau calme. Bien que la végétation aquatique ne semble pas avoir d’incidence sur la présence de ces anisoptères, ces biotypes ont en commun une végétation terrestre très dense. En fin de journée, ces libellules peuvent ainsi se poser sur certains arbustes (Kalmia angustifolia, Viburnum cassinoides, Alnus incana ssp. rugosa) ou sur des branches et des troncs d’arbre ensoleillés (Savard, 2013).
IV- Alimentation
Les larves de Sympetrum vicinum se trouvent au fond des lacs et des étangs. Elles attaquent les proies qui passent à proximité. Les adultes, quant à eux, se posent sur la végétation et attendent que de petits insectes volants passent près d’eux. C’est alors qu’ils s’envolent et partent à la poursuite de leur proie (Corbet, 1999). Préférant des insectes d’une grandeur de 2mm, ces libellules peuvent toutefois se rabattre sur de plus grands spécimens (3-5 mm) lorsque la rigueur de l’automne affecte la population de proies
V- Cycle de vie
Les Odonates ont une métamorphose incomplète (hémimétabole) : leur cycle vital se fait en trois étapes différentes : l’œuf, la larve et l’adulte. La durée de vie d’un sympétrum tardif n’excède pas un an.
Les oeufs éclosent au début du printemps quand la température de l’eau atteint 10°C. Le stade larvaire n’a lieu qu’au printemps et à l’automne. Après la mue, la larve grandit et change de couleur. Ce processus dure environ une heure. Après plusieurs mues, Sympetrum vicinum montre des signes de sa forme finale. Un individu met entre une et sept semaines avant d’atteindre la forme adulte à partir du début du mois de juin. Dès ce moment, la libellule n’a que deux buts : manger et se reproduire. Le stade pré-reproductif (ténéral) peut durer entre trente et quatre-vingt-sept jours selon la zone où on trouve les individus. Une fois la maturité sexuelle atteinte, les libellules cherchent un partenaire, pondent des oeufs puis meurent peu de temps après. (Corbet, 1999)
L’adulte de Sympetrum vicinum a une période d’activité de vol parmi les plus tardives, débutant au plus tôt à la fin juillet et pouvant s’étirer jusqu’en début novembre. À la fin de cette période, l’accouplement et la ponte peuvent encore avoir lieu dans certaines flaques d’eau ensoleillées où le vent est doux, alors que d’autres ont déjà commencé à geler (Perron et al, 2007). Cette tardiveté est permise par des adaptations physiologiques telles que la faible température minimale nécessaire à leur vol ainsi que leur digestion plutôt rapide à température basse. De plus, un postural leur permet de conserver une température corporelle stable jusqu’à un minimum de 10 degrés Celsius (May, 1998).
VI- Reproduction
La fécondation se fait en vol après que le mâle ait identifié une femelle de la même espèce. Le sperme est alors inséré dans la femelle. Après la fécondation, le mâle et la femelle demeurent attachés ensemble en tandem. Lorsqu’elle est à la recherche d’un lieu de ponte, une paire de libellules peut être attirée par la présence d’autres paires, suggérant une attraction mutuelle entre elles qui n’affecte pas l’efficacité de la ponte. Cette présence témoigne également de l’absence de prédateurs dans les environs (McMillan, 2000). La ponte se fait par la femelle lorsqu’elle trempe la pointe de son abdomen à la surface d’un plan d’eau. Ce n’est qu’à ce moment que les deux pièces génitales se détachent l’un de l’autre (Corbet, 1999).
VII- Écologie
Selon une étude effectuée en 2013 sur les odonates du Lac-Saint-Jean, la population de Sympétrum tardif située la plus au nord a été répertoriée dans le parc national de la Pointe-Taillon (PNPT). Ces chercheurs ont avancé que son aire de répartition devait être plus étendue il y a 6000 à 7000 ans, alors que le climat était plus clément. Le refroidissement de la région ainsi que la formation de tourbières acides auraient causé un isolement de la population. C’est dans l’habitat du castor qu’elle a toutefois trouvé refuge, puisque ce mammifère «régule le débit des cours d’eau, tamponne les apports acides, augmente la température de l’eau en aval des étangs, éclaircit le couvert forestier et crée une variété d’habitats humides» (Savard, 2013). Cette coexistence a donc été primordiale à la survie des Sympetrum vicinum dans la région.
Bibliographie
Corbet, P. (1999). Dragonflies : Behavior and Ecology of Odonata. Ithaca, New York: Cornell University Press.
May, M. L. (1998). Body temperature regulation in a late-season dragonfly, Sympetrum vicinum (Odonata: Libellulidae). Dans International Journal of Odonatology (Vol. 1, p.1-13). New Jersey : Taylor & Francis Group.
McMillan, V. E. (2000). Aggregating Behavior During Oviposition in the Dragonfly Sympetrum vicinum (Hagen) (Odonata: Libellulidae). Dans The American Midland Naturalist (Vol. 144, p. 11-18). New York: University of Notre Dame.
Perron, J.-M. & Ruel, Y. (2007). Saison de vol des Odonates du Territoire du marais Léon-Provancher, Neuville, division de rencensement de Portneuf (Québec). Dans Le Naturaliste canadien (Vol. 126, n.2, p.13-17). Québec : GID.
Perron, J.-M., Jobin, L.-J. & Mochon, A. (2005). Odonatofaune du parc national de la Yamaska, division de recensement de Shefford, Québec. Dans Le Naturaliste canadien (Vol 129, n.2, p.17-25). Société Provancher d’histoire naturelle du Canada.
Robert, A. (1963). Les libellules du Québec. Station biologique du Mont Tremblant, Service de la recherche, Ministère de la chasse et des pêcheries.
Savard, M. (2013). Inventaire automnal des odonates au Saguenay-Lac-Saint-Jean : découverte d’une population du sympétrum tardif. Dans Le Naturaliste canadien (Vol. 137, n.1, p.25-32). Société Provancher d’histoire naturelle du Canada.
Worthington, A., Haggert, K. & Loosemore, M. (2005). Seasonality of prey size selection in adult Sympetrum vicinum (Odonata : Libellulidae). Dans International Journal of Odonatology (Vol.8, n.1, p.169-176). NY: Taylor & Francis Group.
Où se situe cette mystérieuse Aeshna eremita dans la classification?
On connaît tous très bien les libellules et les demoiselles, mais on ne connaît pas vraiment les caractéristiques qui les différencient entre elles. En effet, on retrouve dans l’ordre des Odonates ces deux sous-ordres bien distincts : les anisoptères et les zygoptères. On les dissocie entre autres par leurs ailes. Les zygoptères possèdent des ailes antérieures et postérieures identiques qui se replient l’une contre l’autre au repos. Au contraire, les anisoptères possèdent des ailes antérieures et postérieures asymétriques : leurs ailes postérieures sont plus larges à la base que leurs ailes antérieures (2). De plus, lorsqu’au repos, ces ailes sont disposées à plat de chaque côté de l’insecte. Comme on peut le voir sur la Figure 1, notre insecte mystère est bel et bien du sous-ordre des anisoptères. Pour ce qui est de la famille Aeshnidae, elle englobe des espèces de grande taille soit de plus de 70 mm qui ont un vol puissant et qui possèdent des yeux qui se touchent sur une bonne distance sur le dessus de la tête, comme le démontre la Figure 4 (3). Cette famille se caractérise également par la position de certaines parties des ailes. En effet, tel qu’illustrés dans la Figure 2, les triangles des ailes antérieures et postérieures sont placés à égale distance de l’arculus et ils sont orientés vers l’apex (2). Finalement, c’est dans cette famille qu’on retrouve le genre Aeshna qui se distingue des autres par la présence d’une double ramification de la cinquième nervure costale radiale donc au niveau du haut et de l’apex de l’aile (Figure 2) (2). L’espèce Aeshna eremita se trouve donc à cet endroit dans la classification.
Aeshna eremita est la plus grosse espèce du genre Aeshna : les mâles peuvent atteindre une longueur de 75 mm et les femelles de 73 mm (4). De plus, les ailes ont toujours une longueur supérieure à 40 mm (2). Ces dernières se distinguent bien des autres par la position du début de la nervure radiale postérieure supplémentaire qui est sous le premier tiers du ptérostigma (2). Cette nervure est représentée en orange sur la Figure 3.
Aeshna eremita possède une bande noire, épaisse et horizontale sur son front appelée bande frontoclypéale présentée à la Figure 4 (2). De plus, deux caractéristiques la différencient de Aeshna canadensis, son visage de couleur jaune-vert et son absence de points sur l’abdomen (4).
D’autres bandes lui sont également caractéristiques comme celles sur son thorax qui ont une couleur bleu-verdâtre (5). La première bande mésothoracique antérieure est complète. Les autres bandes du ptérothorax sont relativement larges et sont aussi complètes. Il peut également y avoir de petites taches plus pâles entre les bandes thoraciques (Figure 5) (2).
Quand, où et dans quels milieux la retrouve-t-on ?
Aeshna eremita se retrouve dans une grande partie de l’Amérique du Nord. Elle occupe d’est en ouest le Canada, de la partie est de l’Alaska à Terre-Neuve et auLabrador. Au sud, plusieurs états du Nord des États-unis limitent son aire de répartition : Minnesota (6), Michigan, Washington, Dakota du Nord, Utah, Colorado (4) et New-York (6). On la retrouve donc davantage dans les régions boréales (4). Au Québec, on peut rencontrer cette magnifique espèce aussi bien dans les Laurentides qu’au Lac St-Jean ou en Abitibi (5). C’est d’ailleurs l’espèce d’Aeshna qui s’adapte le mieux à tous les types de lacs (5). Elle se retrouve fréquemment près des lacs boisés, des étangs, des marais, des tourbières, des cours d’eau lents et où la végétation est plutôt clairsemée (6). Au repos, cette espèce adopte une position verticale et se pose sur les troncs d’arbres ou les branches de préférence (6). On peut bien les observer au lever et au coucher du soleil, quand la lumière est plutôt faible et que les températures sont plus froides (4). Pour ce qui est de sa métamorphose, elle débute généralement vers la mi-juin (4). C’est vers la mi-juillet que les femelles commencent à pondre et que les mâles recherchent activement à se reproduire (5). On les voit voler jusqu’à la fin octobre, mais la période où elles sont en plus grand nombre est au début du mois juillet jusqu’au début du mois de septembre (4).
Comment et de quoi Aeshna eremita se nourrit-elle ?
Un fait intéressant chez les libellules, y compris celles du genre Aeshna, c’est le mode d’alimentation des larves. Les larves sont carnivores, elles se servent de leur labium modifié pour capturer des proies. Le labium est replié sous la tête de la larve et lorsqu’une proie passe, elle peut projeter très rapidement cette pièce buccale pour l’attraper (7). Ses palpes labiaux en forme de pince l’aide grandement dans sa quête de nourriture (http://www.youtube.com/watch?v=g7Q0IXBM-4M). Chez les larves de la famille des Aeshnidae, les palpes sont courts et précis (7). Tout comme les autres libellules, les espèces du genre Aeshna sont des prédatrices et capturent leurs proies en plein vol. Leur thorax est dirigé vers l’avant permettant une meilleure capture des proies à l’aide des leurs pattes (Figure 6).
Elles se nourrissent principalement de petits diptères et de moustiques (8) de même que de lépidoptères et d’éphéméroptères (5). Les Aeshnidae se nourrissent aussi de proies plus volumineuses comme de trichoptères et même d’autres libellules ou demoiselles. Étant donné la nature de leurs proies, les libellules semblent intéressantes pour la lutte contre des espèces nuisibles (8).
Quel est le cycle de vie de cette belle libellule ?
Les mâles de l’espèce A.eremita volent rapidement au-dessus du littoral et restent habituellement au-dessus ou près de l’eau. En effet, ils vont dans la végétation dense seulement lorsqu’ils sont à la recherche de femelles (6). Au bout de l’abdomen, les mâles ont des appendices anaux qui sont constitués d’une paire de cerques et d’un épiprocte (Figure 7).
Ces appendices servent à attraper la femelle derrière la tête lors de l’accouplement. Le mâle produit du sperme à partir de glandes près de l’extrémité de son abdomen, il doit ensuite transférer le sperme dans un organe accessoire situé sur la face inférieure des segments 2 et 3. Les femelles, de leur côté, portent leurs organes génitaux sur la face inférieure des segments 8 et 9 (Figure 8)
Les femelles peuvent être lignées bleues ou vertes et elles ont des cerques relativement larges comparativement aux mâles. Les femelles ont de larges ovipositeurs qui sont constitués de trois paires de valves dentelées utilisées pour faire des incisions dans le tissu des plantes pour y pondre les œufs (9). Elles pondent généralement sur des morceaux de bois flottant à la surface de l’eau, sur des tiges émergentes de plantes aquatiques ou directement sous la surface de l’eau (6). Les larves sont aquatiques et robustes, elles respirent par des branchies situées à l’intérieur de l’abdomen. De plus, elles se nourrissent d’insectes et parfois de petits poissons. À l’éclosion des œufs, les larves sortent et grandissent progressivement en changeant d’exosquelette (mues) afin remplacer celui qui est trop petit par un nouveau plus grand. Le stade larvaire peut durer plusieurs années. Quand celles-ci arrivent au dernier stade de mue, elles entreprennent leur métamorphose (9), vers la mi-juin habituellement (5) (https://www.youtube.com/watch?v=wUvv4sJnMek). Pendant cette métamorphose, elles vont développer tous les organes nécessaires à la vie adulte sur terre et dans les airs et c’est lorsque le développement est à point qu’elles s’accrochent à une tige de plante et émergent. Elles sortent alors de leur exuvie (exosquelette larvaire) sous la forme adulte. C’est un moment critique pour l’insecte, son corps et ses ailes sèchent pour devenir rigides, il est donc très vulnérable aux prédateurs. La vie d’adulte ne dure que quelques semaines, le but ultime étant la reproduction. L’insecte se nourrit alors sans arrêt jusqu’à l’atteinte de la maturité sexuelle puis s’accouple pour poursuivre son cycle de vie (9).
Le spécimen a été capturé à la Station de
Biologie des Laurentides le 6 Septembre 2013
Classification Règne Animalia Embranchement Arthropoda Classe Insecta Sous-classe Pterygota Division Palaeoptera Ordre Odonata Sous-ordre Anisoptera Famille Aeshnidae GenreAeshna Espèce Aeshna constricta
Les Odonates, terme dérivé du nom scientifique Odonata crée par Fabricius en 1793, signifie en grec « mandibules munies de dents » et désigne l’ordre des Libellules qui représente pour les naturalistes, un important groupe d’insectes à découvrir. Ils sont souvent étudiés dans leur milieu naturel durant le jour ou au crépuscule et sont faciles à observer en raison de leur grande taille et leur corps très colorés et protubérants. Au Québec, on retrouve deux sous-ordres des Odonata, soit les Zygoptera et les Anisoptera qui signifient respectivement « joindre des ailes » et « ailes inégales » (Pilon et Lagacé, 1998). Parmi les Anisoptera, on retrouve la famille des Aeshnidae, le genre Aeshna et l’espèce qu’on étudie Aeshna constricta Say, 1839.
Biologie générale
L’abondance des odonates sur les divers territoires varie en fonction de leur comportement, leur biologie ainsi que leur saison de vol (Perron et al.). Groupe d’insectes prédateurs autant au stade larvaire qu’adultes, ils représentent un élément important de la chaine alimentaire. Ils sont carnivores et attrapent leurs proies au-dessus des étangs, des lacs et de ruisseaux (Wisconsin Aquatic and Terrestrial Resources Inventory). Aeshna constricta est un des grands prédateurs aquatiques avec les espèces Anax junius et Aeshna interrupta (Schultz, 2009). Dû à leur diversité et leur nombre dans certaines régions, ils accomplissent leur cycle vital dans les régions humides semi-inondées et sont indicateurs de l’excellence de la qualité biologique du milieu. Les odonates à l’état larvaire durent plus longtemps que l’état adulte et leur métamorphose se fait en une saison, mais des fois prennent plusieurs années, dépendamment de l’espèce et de la température des eaux. Les deux espèces Aeshna constricta et Aeshna interrupta passent deux ou plusieurs hivers au stade larvaire et nécessitent donc des habitats dans des marécages profonds et régulièrement inondés (Schultz, 2009). Plus particulièrement, les recherches effectuées au Québec par Pilon et Lagacé (1998) ont démontré une saison de vols spécifique pour les diverses espèces odonates, d’où la probabilité de les capturer à diverses périodes. En général, la saison s’étend de la mi-mai à la mi-novembre, mais contrairement à la généralité, Aeshna constricta est une des espèces qui présente une activité de vol plus longue à l’automne qui s’étend du mois d’août jusqu’au début du mois d’octobre (Perron et Ruel, 2002)(Voir tableau II). Enfin une des particularités physiologiques des femelles de l’espèce Aeshna constricta est qu’elles possèdent un ovipositeur aidant à pondre leurs œufs sur des plantes aquatiques (Cannings, 2002).
La distribution de l’espèce
L’hémisphère nord de la Terre se divise en une région arctique et une région subarctique qui se subdivise en une zone paléarctique et une zone néarctique. Cette dernière appartient à la région arctique qu’on nomme « Le Nouveau-Monde » soit l’Amérique du Nord où l’on retrouve une petite diversité d’Odonate. En effet, parmi les espèces habitant cette zone tempérée froide (holarctique) l’espèce Aeshna constricta a été retrouvée au Canada (Québec, Île-du-Prince-Édouard) et aux États-Unis (Utah, Illinois, Connecticut, New Hampshire) (Figure 1). Cette espèce fréquente plusieurs biotopes, mais surtout des étangs (Pilon et Lagacé, 1998) et des marais (Hilton, 1987). Une étude approfondie sur l’odonatofaune du parc national de la Yamaska a été réalisée entre 2002 et 2004, du printemps jusqu’à l’automne, afin de répertorier la grande diversité des odonates dans un espace aussi restreint dépendamment de leur saison de vol. Suite aux résultats, Aeshna constricta a été retrouvée dans le réservoir Choinière de la rivière Yamaska Nord, dans l’étang à castor présentant une surface d’eau suffisamment dégagée et dans les marais au nord du réservoir dans les milieux forestiers mixtes (Perron et al.). L’espèce a également été retrouvée dans quelques secteurs du sud du Québec, soit dans la région du Mont Pinacle (Legault, 1975 et 1977) et dans le secteur de Granby au Lac Boivin (Legault, 1979) ; mais leur diversité reste tout de même inférieure à celle présente au parc national de la Yamaska. Les recherches effectuées par l’entomologiste Raymond Hutchison entre les mois de juillet à septembre ont également signalé la présence de plusieurs espèces d’Aeshna à l’état adulte près des rives de l’Outaouais (Hutchison, 2011).
la morphologie
L’espèce Aeshna constricta a une taille qui varie en moyenne entre 2,7 et 2,8 pouces (Wisconsin Aquatic and Terrestrial Resources Inventory). Elle possède en général un thorax brun ou noir avec des tâches brunes, vertes ou jaunes et des segments abdominaux avec une seule carène de chaque côté. Le prothorax est muni de deux bandes latérales dont la première est large et non recourbée deux fois, alors que la bande postérieure présente une projection vers le bas dans la partie supérieur (Pilon et Lagacé, 1998). Souvent, on les différencie de l’espèce Aeshna canadensis par l’absence d’une tache jaune arrondie et bien délimitée entre les deux bandes latérales. Les taches abdominales chez les mâles sont typiquement bleues alors que chez les femelles les taches peuvent être bleues, vertes, ou jaunes. Contrairement aux mâles, les femelles ont le 9e segment de l’abdomen plus large que le 8e (Wisconsin Aquatic and Terrestrial Resources Inventory). De plus la tête, de couleur verte pâle, est globuleuse et possède des yeux composés contigus. Le front porte distinctivement une marque en forme de T, autrement nommé le «T-spot» (Cannings, 2002). La bande frontoclypéale (suture en dessous du front) est mince et brunâtre ou de même couleur que le front et que le clypeus.
Aeshna constricta possède deux paires d’ailes de longueur supérieure à 40 mm qui sont rattachées au mésothorax et au métathorax dont chacune possède un stigmate. Une des façons de les distinguer des espèces de Zygoptera est d’analyser la forme des ailes antérieures et postérieures qui sont de forme nettement différente chez les Anisoptera (antérieur supérieur à postérieur) (Pilon et Lagacé, 1998).
Hilton, D.F.J. 1987. Aquatic insects of peatlands and marshes in Canada. Memoirs of the Entomological Society of Canada 140: 57-63.
Hutchinson, R. 2001. Les libellules (Odonata) des grandes rivières du sud du Québec, Biodiversity,2:2, 9-16, [En ligne]. http://dx.doi.org/10.1080/14888386.2001.9712542 (consulté le 27 octobre 2013)
Legault, J. 1975. Les Odonates de la région du mont Pinacle, sud du Québec. Cordulia, 1 : 121-123
Legault, J. 1977. Liste de contrôle préliminaire des Odonates de la région du mont Pinacle, sud du Québec. Cordulia, 3 : 76-78
Legault, J. 1979. Liste préliminaire des Odonates du lac Boivin, Granby (Shefford), Québec. Fabreries, 6 : 73
Pilon, J.-G. et D. Lagacé, 1998. Les Odonates du Québec. Entomofaune du Québec (EQ) inc., Chicoutimi, Québec. 367p
Perron, J.M, Jobin, L.-J. et Mochon, A. 2005. Odonatofaune du parc national de la Yamaska, division de recensement de Shefford, Québec. Le Naturaliste canadien, 129 (2) : 17-25 [En ligne]. http://www.provancher.qc.ca/upload/file/129_2%20p%2017-25.pdf (consulté le 25 octobre 2013)
Perron, J.M et Y.Ruel. 2002. Saison de vol des Odonates du territoire du marais Léon-Provancher, Neuville, division de recensement de Portneuf (Québec). Le Naturaliste canadien, 126 (2) : 13-17 [En ligne]. http://www.provancher.qc.ca/upload/file/126_2%20p%2013-17.pdf (consulté le 26 octobre 2013)
Schultz, T.D. 2009 Diversity and Habitats of a Prairie Assemblage of Odonata at Lostwood National Wildlife Refuge, North Dakota. Journal of the Kansas Entomological Society 82(1):91-102. 2009 [En ligne]. http://dx.doi.org/10.2317/JKES803.24.1 (consulté le 25 octobre 2013)
U.S. States and Canadian Provinces – Aeshna constricta. Encyclopedia of Life. [En ligne]. <http://eol.org/data_objects/14877572>. (consulté le 27 octobre 2013)















.jpg)





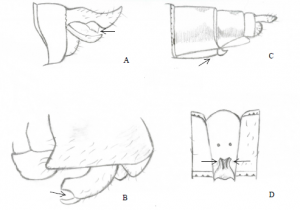






















 L’hémisphère nord de la Terre se divise en une région arctique et une région subarctique qui se subdivise en une zone paléarctique et une zone néarctique. Cette dernière appartient à la région arctique qu’on nomme « Le Nouveau-Monde » soit l’Amérique du Nord où l’on retrouve une petite diversité d’Odonate. En effet, parmi les espèces habitant cette zone tempérée froide (holarctique) l’espèce Aeshna constricta a été retrouvée au Canada (Québec, Île-du-Prince-Édouard) et aux États-Unis (Utah, Illinois, Connecticut, New Hampshire) (Figure 1). Cette espèce fréquente plusieurs biotopes, mais surtout des étangs (Pilon et Lagacé, 1998) et des marais (Hilton, 1987). Une étude approfondie sur l’odonatofaune du parc national de la Yamaska a été réalisée entre 2002 et 2004, du printemps jusqu’à l’automne, afin de répertorier la grande diversité des odonates dans un espace aussi restreint dépendamment de leur saison de vol. Suite aux résultats, Aeshna constricta a été retrouvée dans le réservoir Choinière de la rivière Yamaska Nord, dans l’étang à castor présentant une surface d’eau suffisamment dégagée et dans les marais au nord du réservoir dans les milieux forestiers mixtes (Perron et al.). L’espèce a également été retrouvée dans quelques secteurs du sud du Québec, soit dans la région du Mont Pinacle (Legault, 1975 et 1977) et dans le secteur de Granby au Lac Boivin (Legault, 1979) ; mais leur diversité reste tout de même inférieure à celle présente au parc national de la Yamaska. Les recherches effectuées par l’entomologiste Raymond Hutchison entre les mois de juillet à septembre ont également signalé la présence de plusieurs espèces d’Aeshna à l’état adulte près des rives de l’Outaouais (Hutchison, 2011).
L’hémisphère nord de la Terre se divise en une région arctique et une région subarctique qui se subdivise en une zone paléarctique et une zone néarctique. Cette dernière appartient à la région arctique qu’on nomme « Le Nouveau-Monde » soit l’Amérique du Nord où l’on retrouve une petite diversité d’Odonate. En effet, parmi les espèces habitant cette zone tempérée froide (holarctique) l’espèce Aeshna constricta a été retrouvée au Canada (Québec, Île-du-Prince-Édouard) et aux États-Unis (Utah, Illinois, Connecticut, New Hampshire) (Figure 1). Cette espèce fréquente plusieurs biotopes, mais surtout des étangs (Pilon et Lagacé, 1998) et des marais (Hilton, 1987). Une étude approfondie sur l’odonatofaune du parc national de la Yamaska a été réalisée entre 2002 et 2004, du printemps jusqu’à l’automne, afin de répertorier la grande diversité des odonates dans un espace aussi restreint dépendamment de leur saison de vol. Suite aux résultats, Aeshna constricta a été retrouvée dans le réservoir Choinière de la rivière Yamaska Nord, dans l’étang à castor présentant une surface d’eau suffisamment dégagée et dans les marais au nord du réservoir dans les milieux forestiers mixtes (Perron et al.). L’espèce a également été retrouvée dans quelques secteurs du sud du Québec, soit dans la région du Mont Pinacle (Legault, 1975 et 1977) et dans le secteur de Granby au Lac Boivin (Legault, 1979) ; mais leur diversité reste tout de même inférieure à celle présente au parc national de la Yamaska. Les recherches effectuées par l’entomologiste Raymond Hutchison entre les mois de juillet à septembre ont également signalé la présence de plusieurs espèces d’Aeshna à l’état adulte près des rives de l’Outaouais (Hutchison, 2011). L’espèce Aeshna constricta a une taille qui varie en moyenne entre 2,7 et 2,8 pouces (Wisconsin Aquatic and Terrestrial Resources Inventory). Elle possède en général un thorax brun ou noir avec des tâches brunes, vertes ou jaunes et des segments abdominaux avec une seule carène de chaque côté. Le prothorax est muni de deux bandes latérales dont la première est large et non recourbée deux fois, alors que la bande postérieure présente une projection vers le bas dans la partie supérieur (Pilon et Lagacé, 1998). Souvent, on les différencie de l’espèce Aeshna canadensis par l’absence d’une tache jaune arrondie et bien délimitée entre les deux bandes latérales. Les taches abdominales chez les mâles sont typiquement bleues alors que chez les femelles les taches peuvent être bleues, vertes, ou jaunes. Contrairement aux mâles, les femelles ont le 9e segment de l’abdomen plus large que le 8e (Wisconsin Aquatic and Terrestrial Resources Inventory). De plus la tête, de couleur verte pâle, est globuleuse et possède des yeux composés contigus. Le front porte distinctivement une marque en forme de T, autrement nommé le «T-spot» (Cannings, 2002). La bande frontoclypéale (suture en dessous du front) est mince et brunâtre ou de même couleur que le front et que le clypeus.
L’espèce Aeshna constricta a une taille qui varie en moyenne entre 2,7 et 2,8 pouces (Wisconsin Aquatic and Terrestrial Resources Inventory). Elle possède en général un thorax brun ou noir avec des tâches brunes, vertes ou jaunes et des segments abdominaux avec une seule carène de chaque côté. Le prothorax est muni de deux bandes latérales dont la première est large et non recourbée deux fois, alors que la bande postérieure présente une projection vers le bas dans la partie supérieur (Pilon et Lagacé, 1998). Souvent, on les différencie de l’espèce Aeshna canadensis par l’absence d’une tache jaune arrondie et bien délimitée entre les deux bandes latérales. Les taches abdominales chez les mâles sont typiquement bleues alors que chez les femelles les taches peuvent être bleues, vertes, ou jaunes. Contrairement aux mâles, les femelles ont le 9e segment de l’abdomen plus large que le 8e (Wisconsin Aquatic and Terrestrial Resources Inventory). De plus la tête, de couleur verte pâle, est globuleuse et possède des yeux composés contigus. Le front porte distinctivement une marque en forme de T, autrement nommé le «T-spot» (Cannings, 2002). La bande frontoclypéale (suture en dessous du front) est mince et brunâtre ou de même couleur que le front et que le clypeus. Aeshna constricta possède deux paires d’ailes de longueur supérieure à 40 mm qui sont rattachées au mésothorax et au métathorax dont chacune possède un stigmate. Une des façons de les distinguer des espèces de Zygoptera est d’analyser la forme des ailes antérieures et postérieures qui sont de forme nettement différente chez les Anisoptera (antérieur supérieur à postérieur) (Pilon et Lagacé, 1998).
Aeshna constricta possède deux paires d’ailes de longueur supérieure à 40 mm qui sont rattachées au mésothorax et au métathorax dont chacune possède un stigmate. Une des façons de les distinguer des espèces de Zygoptera est d’analyser la forme des ailes antérieures et postérieures qui sont de forme nettement différente chez les Anisoptera (antérieur supérieur à postérieur) (Pilon et Lagacé, 1998).