Par Delphine EYMIEU et Lucille JOYEUX
Texte et figures (sauf si indiqué) CC BY-SA 2018, par les auteures.
Anax junius (Drury, 1773) l’anax précoce
L’espèce Anax junius a été décrite pour la première fois en 1773 par Drury. Notre spécimen (Figure 1) a été attrapé le 06 septembre 2018 à l’aide d’un filet fauchoir sur le Chemin du Lac Croche menant à la station de biologie des Laurentides (Figure 2) aux coordonnées suivantes: Latitude 45.9842 | Longitude -74.009


Classification
Embranchement Arthropoda
Sous-embranchement Hexapoda
Classe Insecta
Ordre Odonata
Sous-ordre Anisoptera
Famille Aeshnidae
Genre Anax
Espèce Anax junius
Identification
Pour identifier précisément notre spécimen, nous avons utilisé deux clés d’identification (Robert, 1957 ; Pilon et Lagacé, 1998). Le processus d’identification de notre spécimen a été relativement simple grâce à sa grande taille (7.4cm) qui a facilité nos observations.
Nous avons d’abord chercher la famille de notre odonate grâce à la clé d’identification des familles de Robert A. (1957).
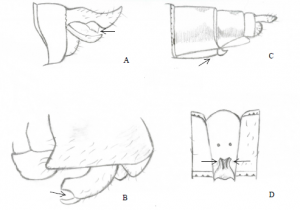
On remarque que la largeur de la base des ailes est différente, en effet, la base de l’aile postérieure est plus large que la base de l’aile antérieure. Ce caractère nous a permis d’approuver que notre spécimen provient du sous-ordre Anisoptera, soit plus communément des libellules. Ensuite, on voit que la distance entre l’arculus et le triangle est à peu près égale dans les deux paires d’ailes (Figure 3). Le ptérostigma, qui est la tâche noire à l’extrémité de chaque aile, est pourvu d’une grosse nervure oblique à son bord interne, et on observe également que les yeux se touchent. Ces 3 observations de caractères permettent de dire que notre anisoptère est plus précisément de la famille des Aeshnidae.

Une fois notre famille trouvée, nous avons cherché l’espèce grâce à la clé d’identification de Pilon J-G. et Lagacé D. (1998).
Cette recherche a été très simple puisque notre espèce a un caractère bien précis qui la différencie des autres Aeshnides de l’Amérique du Nord : son thorax de couleur vert. Ce caractère est unique à l’espèce Anax junius en Amérique du Nord.
Nous avons également pu déterminer facilement le sexe de notre Anax junius puisque cette espèce a un dimorphisme sexuel visuel : la couleur de son abdomen. Chez les mâles, l’abdomen est de couleur bleue tandis que chez la femelle, l’abdomen est de couleur brune. Notre Anax junius est donc une femelle.
Description
Anax junius fait partie des plus grosses libellules, atteignant 7 ou 8 cm de long. Leurs yeux sont grands, leur thorax est court et robuste et leur abdomen est long et mince. Tous les individus, mâles et femelles, possèdent un thorax vert et une ligne noire qui s’étend sur leur surface supérieure. (Needham et Westfall, 1955). Cependant, les individus sont polymorphes selon le sexe. Les mâles ont le thorax, le visage, et les yeux de couleur verts ternes. Le premier segment de leur abdomen est vert, et les 2 à 6 autres segments sont de couleurs bleu vif après quoi l’abdomen s’estompe en vert terne puis devient sombre. Chez la plupart des femelles, les yeux sont brun, les deux premiers segments abdominaux sont verts, et le reste de l’abdomen est brun rougeâtre (Fraker et Luttbeg, 2012). Chez le juvénile, l’abdomen est vert terne, rougeatre ou brun. Les ailes sont incolores ou orange et prennent une couleur ambrée à mesure qu’elles vieillissent, notamment chez les femelles. (Fraker et Luttbeg, 2012)
Cycle de vie
Les libellules Anax junius sont hémimétaboles, c’est-à-dire sans stade immobile entre la larve et l’adulte.
1. La reproduction a normalement lieu dans un habitat d’eau calme, par exemple les étangs, les lacs, les marais, les marécages, les cours d’eau lents. (Beaton, 2007).
2. Les œufs sont pondus dans des habitats aquatiques et éclosent après au moins 6 ou 7 jours. Les œufs éclosent en nymphes et subissent 9 à 13 instars nymphaux. Les nymphes ont un abdomen ovale allongé, de grands yeux et un grand labium qui peut être étendu pour capturer une proie. Les nymphes Anax junius ont des labiums plats avec des palpes pointues qui sont utilisées pour lancer une proie. Ils sont généralement de couleur marron ou verte. Les derniers stades peuvent atteindre 3,5 à 5 cm de long (Paulson, 2011).
3. Lorsqu’elles atteignent la fin du stade nymphal, les nymphes sortent de l’eau et cherchent un endroit pour émerger de leur exuvie. Leurs ailes se déploient à mesure que les ailes et le corps se dilatent. À ce moment là, elles sont des adultes sexuellement immatures. Elles quittent leur peau nymphale et gagnent rapidement en masse. La cuticule durcit et leur couleur devient plus distinct. Le temps total de développement des nymphes peut prendre presque 1 an. Les nymphes terminent leur développement à l’âge adulte en juin et en juillet. (Hopkins et al, 2011)
4. Les adultes ne vivent que plusieurs semaines à un peu plus d’un mois après avoir atteint l’âge adulte (Paulson, 2011).
Distribution géographique
Anax Junius est répandue majoritairement dans la région du Néarctique. Elles sont présentes sur l’ensemble des États-Unis, du Canada et du Mexique (Figure 4).
Leur aire de répartition s’étend également à Hawaii, à Tahiti, aux Antilles, en Chine et dans la péninsule en Russie. Parfois les individus sont emportés par des tempêtes ou des vents violents dans des régions comme aux Bermudes, au Royaume-Uni et en France. Anax Junius reste l’une des libellules les plus répandues en Amérique du Nord. (Corbet, 1999)

Populations de Anax junius
Cette espèce a deux types de population différentes: les populations résidentes et migratrices.
Les populations résidentes restent dans la zone générale d’où elles émergent. Pour les populations résidentes du Nord, les adultes s’accouplent et pondent de fin juillet à août. La progéniture qui en résulte éclot et se développe vers le mi-stade puis hiverne lorsque la température baisse. Le temps total de développement des nymphes peut prendre de 11 à 12 mois pour les résidentes, puisque les nymphes ont terminé leur développement lorsque les températures se réchauffent au printemps, puis apparaissent à l’âge adulte en juin et en juillet (Trottier, 1971).
Les adultes qui migrent vers le Nord tendent à arriver avant l’émergence des résidentes du Nord au printemps. Les adultes migrateurs s’accouplent et pondent en juin. Contrairement aux populations résidentes, le développement nymphal des progénitures migrantes ne prend que 3 à 5 mois et elles ne passent pas l’hiver. Elles émergent généralement en fin août et en septembre. Elles commencent souvent leur migration à l’âge immature, se nourrissant en cours de route pour devenir adultes. Les adultes s’accouplent et pondent pendant la migration et lorsqu’ils atteignent leurs destinations méridionales. La progéniture produite se développe pendant l’hiver chaud austral, avant de devenir adulte et de migrer vers le Nord (Trottier, 1971).
La migration nécessite non seulement des comportements spécifiques au stade adulte mais également des adaptations du développement des larves aquatiques. La prédation des poissons a une grande influence sur la survie et le développement des larves. La survie des larves sont plus élevée si les poissons insectivores sont absents. Les conditions climatiques peuvent favoriser la reproduction et la période de migration. (May et al., 2017)
Bibliographie
Beaton, G. (2007). Dragonflies & Damselflies of Georgia and the Southeast. University of Georgia Press, Athens, Georgia.
Corbet, P. (1999). Dragonflies Behavior and Ecology of Odonata. Ithaca, New York: Cornell University Press
Fraker, M., et B. Luttbeg. (2012). Predator-prey space use and the spatial distribution of predation events. Behaviour, 149(5)
Hopkins, G., Gall, B., et Brodie, E. (2011). Ontogenetic Shift in Efficacy of Antipredator Mechanisms in a Top Aquatic Predator, Anax junius (Odonata: Aeshnidae). Ethology, 117(12)
Needham, J.G. et Westfall, M.J. (1955) The Manual of the Dragonflies of North America (Anisoptera): Including the Greater Antilles and the Provinces of the Mexican Border. University of California Press, Berkeley and Los Angeles.
May, M.L., Gregoire, J.A., Gregoire, S.M., Lubertazzi, M.A., Matthews, J.H. (2017). Emergence phenology, uncertainty, and the evolution of migratory behavior in Anax junius (Odonata: Aeshnidae). PLoS ONE. 12(9)
Paulson, D. (2011). Dragonflies and Damselflies of the East. Princeton, New Jersey: Princeton University Press.
Pilon, J-G., et Lagacé, D. (1998). Les odonates du Québec. Chicoutimi, Canada: Entomofaune du Québec (EQ) Inc.
Robert, A. (1957). Les principales libellules du Québec (Vol. 7, N°9). Ottawa, Canada : Le Jeune Naturaliste.
Trottier, R. (1971). Effect of temperature of the life of Anax junius (Odonata : Aeshnidae) in Canada. The Canadian Entomologist. 103(12)