Par Philippe HÉNAULT et Alexandre HUOT
Texte et figures (sauf si indiqué) CC BY-SA 2018, par les auteurs.

Élytres bruns et bossus qui se fondent bien au travers du bois et de la terre, tendance à faire le mort lorsqu’il se sent menacé… On peut dire que le ténébrion cornu, ou Bolitotherus cornutus, sait se faire discret, et il est très facile de le confondre pour un simple caillou lorsqu’il ne désire pas être vu (comme on peut le voir dans les premières secondes de ce vidéo ). C’était d’ailleurs le cas de notre individu, qui n’était définitivement pas à la vue de tous, mais plutôt installé à l’intérieur même d’un bout de bois mort, sans doute en train d’y manger des champignons.
Classification
Ordre : Coleoptera
Sous-ordre : Polyphaga
Famille : Tenebrionidae
Sous-famille : Bolitophaginae
Tribu : Bolitophagini
Genre : Bolitotherus
Espèce : Bolitotherus cornutus (Panzer, 1794)
Identification
L’identification d’insectes peut s’avérer plutôt ardue, puisqu’il faut utiliser de nombreuses clés d’identifications et regarder des caractères qui sont parfois microscopiques. Heureusement pour nous, Bolitotherus cornutus a une apparence assez caractéristique qui est difficilement oubliée, et nous avions donc une bonne idée de l’espèce dont il s’agissait avant même de commencer l’identification. Malgré tout, nous nous sommes prêtés à l’exercice des clés d’identification afin de connaître les traits, hormis sa forme, qui nous permettait de bien reconnaître cette espèce.

Tout d’abord, l’ordre de l’insecte a facilement été établi par son apparence générale, la partie dure ressemblant à une carapace étant en fait ses élytres (figure 1), caractère propre aux coléoptères. Il a ensuite été possible de déterminer qu’il appartenait au sous-ordre des Polyphaga. La différence entre les Polyphaga et les Adephaga (l’autre grand sous-ordre de coléoptères) réside au niveau des coxas postérieurs et de ce que l’on appelle les sternites, les segments de l’abdomen. Chez les Adephaga, les coxas sont fusionnés aux premiers sternites et les divisent alors que chez les Polyphaga, les coxas sont plutôt situés sur l’épiméron et ne divisent pas le premier sternite (figure 2).

La famille de l’insecte a ensuite été déterminée grâce à une clé d’identification (Choate, 2003) qui nous a menés à la famille des ténébrions (Tenebrionidae). Les traits distinctifs de la clé nous ayant permis d’éliminer les autres familles de Polyphaga sont les élytres couvrant l’abdomen, les tarsomères en formules 5-5-4 (figure 3) (trait qui aura également été utile pour l’identification à la famille, puisqu’il existe des sous-familles de Tenebrionidae qui ont une formule 4-4-4) et les cavités coxales antérieures fermées. Toutefois, les traits les plus importants et distinctifs des ténébrions sont les antennes insérées sous les yeux, de façon à ce que leur insertion ne soit pas visible lorsque l’on voit l’insecte de haut, ainsi que la crête qui sépare une partie des yeux en deux. (figure 4)


Les Tenebrionidae contiennent toutefois plusieurs sous-familles différentes, et il convient donc d’utiliser une nouvelle clé (Choate et al., 2005), plus spécifique, afin de pouvoir identifier notre espèce. Cette nouvelle clé nous a amenés à la sous-famille Bolitophaginae, dans la tribu des Bolitophagini. Les caractéristiques principales de ce groupe sont la présence de griffes simples (figure 4), le tibia des pattes antérieures qui n’est pas denté, le dernier segment de l’antenne qui n’est pas significativement plus long que les autres (figure 5), la membrane visible sur la marge des troisième et quatrième sternites (figure 2) et les « joues » (genae) de l’insecte qui sont dentées (figure 3).
Finalement, il ne reste qu’à faire la distinction entre deux espèces, Bolitotherus cornutus et Bolitotherus corticola. La différence se fait au niveau des antennomères (Choate et al., 2005). Ceux-ci nous ont permis d’identifier avec précision Bolitotherus Cornutus, puisque les antennes étaient composées de dix segments et que le troisième était allongé (figures 5 et 6). Malheureusement, les antennes de notre spécimen étaient tombées, mais il nous a tout de même été possible de compter les dix segments, même s’ils étaient séparés.


Biologie
Comportement, alimentation et reproduction
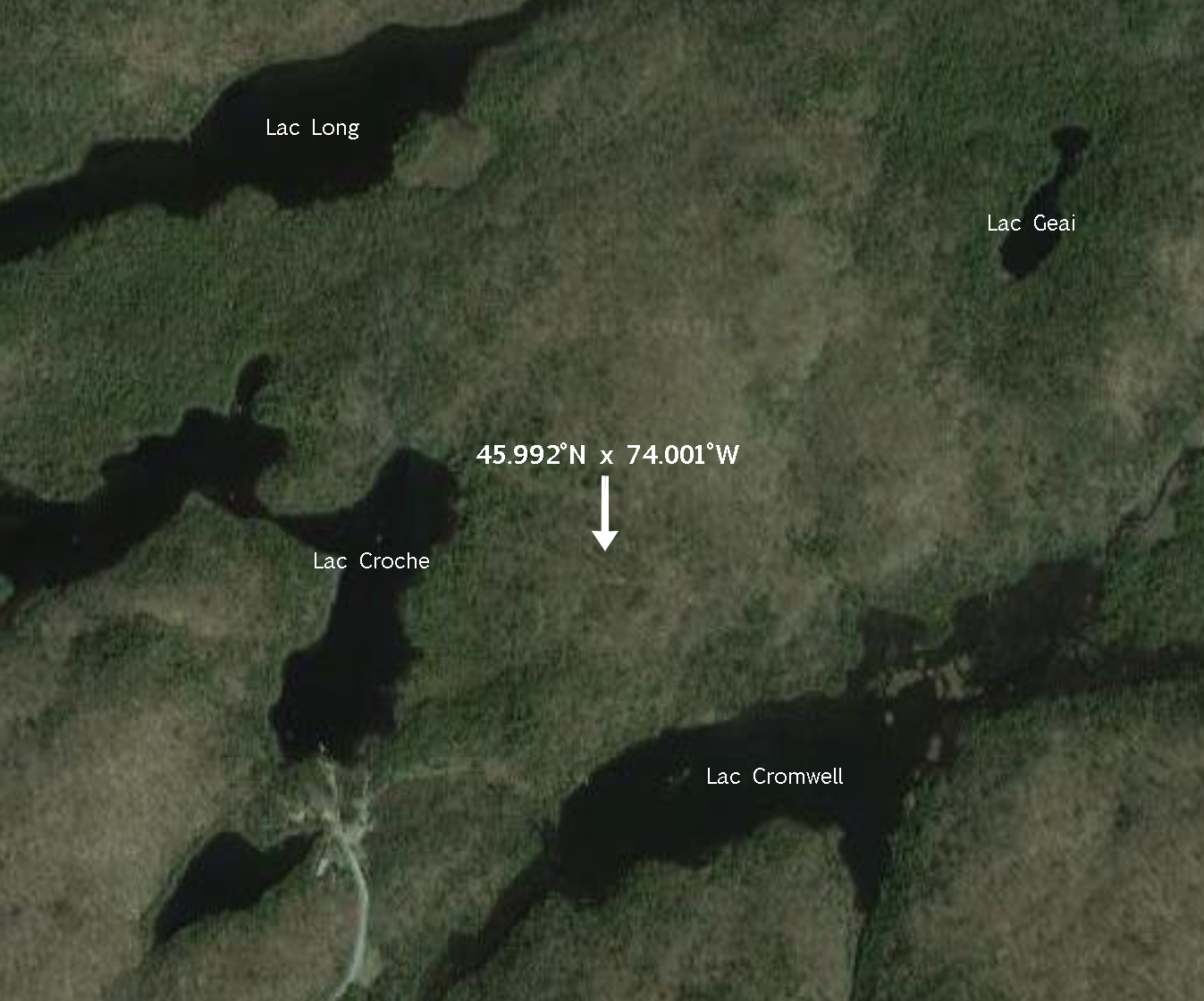
Le ténébrion cornu (Bolitotherus cornutus), vit toute sa vie sur ou près des champignons dit «polypores», qui poussent sur les arbres, ce qui lui donne son nom commun en anglais de «forked fungus beetle» (Liles, 1956). Il se nourrit de la chair du sporophore du champignon hôte tout au long de sa vie (Pace, 1967). Parmi ses hôtes réguliers se retrouvent notamment les champignons Ganodermas applanatum et Ganodermas tsugae, mais il se retrouve sur et se nourrit d’autres espèces de «polypores» (Heatwole, H. & Heatwole, A. , 1968). Il se déplace peu et reste près de l’arbre où poussent les champignons, le vol a cependant été observé en laboratoire (Teichert, 1999). Le Bolitotherus cornutus (Panzer) est nocturne, et peut se cacher dans l’arbre hôte du champignon durant le jour (Liles, 1956), ce qui explique l’emplacement de notre spécimen, qui a été trouvé enfoui dans un arbre mort de la Station de Biologie des Laurentides.
Les combats entre mâles arrivent de façon fréquente. Les mâles peuvent être territoriaux et peuvent se battre pour une ressource commune (Pace, 1967). Cependant, les combats surviennent très souvent lors de l’accouplement et des rituels qui précèdent l’accouplement (Pace, 1967). Les adultes ont en effet un rituel pré-accouplement particulier dans lequel le mâle, placé sur le dos de la femelle, place sa tête vers l’abdomen de la femelle de façon à ce que son abdomen soit vis-à-vis le thorax de celle-ci. Il frotte alors son abdomen contre le thorax de la femelle, ce qui produit un son (Liles, 1956). Si la femelle est réceptive au rituel, le mâle peut ensuite s’accoupler avec elle. Durant le rituel, un autre mâle peut tenter de déloger le mâle en rituel à l’aide de ces paires de cornes. Le mâle qui s’agrippe à la femelle doit donc résister, et la force avec laquelle il tiendra la femelle sera un facteur , et est corrélée avec la taille des pattes et de la taille du mâle qui se défend (Benowitz & al., 2012). La taille des cornes est donc un caractère de sélection sexuelle, puisque celles-ci permettent de mieux déloger les mâles rivaux des femelles (Conner, 1988). Voici un vidéo d’un combat lors d’un rituel précédant un accouplement.
Lorsqu’il se sent attaqué, le ténébrion cornu utilise la technique de beaucoup d’autres ténébrions, soit celle de faire le mort (Liles, 1956). Il peut également émettre une substance au goût et à l’odeur désagréable à partir de ces glandes abdominales (Pace, 1967).
Cycle de vie
Les oeufs sont pondus sur la surface du champignon hôte, souvent en 2 vagues distinctes, une au printemps et une au début de l’automne, dépendant du stade dans lequel émerge le coléoptère à la suite de son hivernation. Les larves et les adultes peuvent hiverner, en continuant le cycle de vie une fois que la température redevient plus clémente (Liles, 1956). Lorsque les oeufs sont pondus, ils sont enduits d’une substance foncée par la femelle, qui durcira par la suite. Cinq jours après l’éclosion des oeufs, qui se passe entre 11 et 26 jours après la ponte, la larve creuse des tunnels dans le champignon hôte et s’en nourrit, elle passe ensuite par 4 à 5 stades larvaires (Liles, 1956). La larve est blanche et segmentée, et a des protubérances sclérifiées en forme de cône sur les mandibules, qui ressortent vers l’avant de sa tête globulaire et plus foncé que le corps (Peterson, 1951). La nymphose se passe dans une chambre à l’intérieur du champignon, où la nymphe est plutôt active (Liles, 1956). Les nymphes mâles et femelles sont distinguables l’une de l’autre puisqu’elles portent les mêmes caractéristiques au niveau de la tête que les adultes, les cornes des mâles et les protubérances de la femelle au niveau de la tête sont donc visibles. Ce stade dure entre 5 et 22 jours. L’adulte émerge par la suite du champignon (Liles, 1956). Voici un vidéo de la nymphe très active du ténébrion cornu.
Références
Benowitz, K. M., Brodie III, E. D., & Vincent A. Formica. (2012). Morphological Correlates of a Combat Performance Trait in the Forked Fungus Beetle, Bolitherus cornutus. PLoS ONE, 7(8), e42738. https://doi.org/10.1371/journal.pone.0042738
Choate, P.M, Dunford, J.C., Thomas, M.C. (2005). The Darkling Beetles of Florida and Eastern United States. Visionné à l’adresse suivante : https://entnemdept.ifas.ufl.edu/teneb/identification.htm
Choate, P. M. (2003). Introduction to the Identification of Beetles (Coleoptera). Visionné à l’adresse suivante : http://www.entnemdept.ufl.edu/choate/beetles1a.pdf
Conner, J. (1988). Field Measurements of Natural and Sexual Selection in the Fungus Beetle, Bolitotherus cornutus. Evolution, 42(4), 736‑749. https://doi.org/10.1111/j.1558-5646.1988.tb02492.x
Heatwole, H., & Heatwole, A. (1968). Movements, Host-Fungus Preferences, and Longevity of Bolitotherus cornutus (Coleoptera: Tenebrionidae). Annals of the Entomological Society of America, 61(1), 18‑23. https://doi.org/doi:10.1093/aesa/61.1.18
Liles, M. P. (1956). A Study of the Life History of the Forked Fungus Beetle, Bolitotherus Cornutus (Panzer) (Coleoptera; Tenebrionidae). Ohio Journal of Science, 56(6), 329‑337. https://kb.osu.edu/bitstream/handle/1811/4397/1/V56N06_329.pdf
Pace, A. E. (1967). Life History and Behavior of a Fungus Beetle Bolitotherus Cornutus (Tenebrionidae). Occasional Papers of the Museum of Zoology, (653), 1‑15. http://hdl.handle.net/2027.42/57089
Peterson, A. (1957). Larvae of Insects, An Introduction to Nearctic Species Part II: Coleoptera, Diptera, Neuroptera, Siphonaptera, Mecoptera, Trichoptera (3rd éd.). Ann Harbor, Michigan: Edward Brothers Inc.







 « Red-lined carrion beetle »
« Red-lined carrion beetle »










































 Bibliographie
Bibliographie


{kind=link}