
– LE FANTÔME DES MARAIS –
Texte, photographies, illustrations et identification du spécimen par
Manon FUSELIER & Mathilde GAUDREAU

Dans la pénombre des marécages de l’est nord-américain, un véritable spectre hante les lieux. Apparenté aux créatures de légendes, son apparition éphémère est un événement auquel seuls les spectateurs les plus attentifs ont la chance d’assister. On le prénomme le fantôme des marais, « phantom crane fly », ou Bittacomorpha clavipes pour les intimes.
Bittacomorpha clavipes est un insecte holométabole dont le nom commun fait référence à sa coloration blafarde, à sa gracilité et à son vol caractéristique qui lui donnent l’apparence d’un véritable spectre livide. Il débute son existence tel un cadavre, sous la forme d’une larve enterrée sous une couche de sédiments organiques humides. Suivant une nymphose souterraine, c’est au stade adulte que la métamorphose se complète et que l’esprit s’extirpe de son linceul pour entreprendre un vol si discret et léger qu’il semble littéralement flotter dans l’air.
Contrairement à ce que son nom commun indique, un spécimen de B. clavipes ne passe pas inaperçu dans une collection en raison de son patron de coloration blanche et noire caractéristique et de la difficulté liée à le préserver intact suivant la capture. En effet, ses longues pattes délicates se détachent aisément; le spécimen présenté fut ainsi collé latéralement sur deux pointes entrecroisées (Figure 1) afin de le préserver dans son intégrité.
CLASSIFICATION
Ordre: Diptera
Sous-Ordre: Nematocera
Super-Famille: Ptychopteroidea
Famille: Ptychopteridae
Sous-Famille: Bittacomorphinae
Genre: Bittacomorpha
Espèce: Bittacomorpha clavipes (Fabricius, 1781)
IDENTIFICATION
Il est plutôt aisé d’identifier précisément un spécimen de B. clavipes car de nombreux caractères clés simples ont été définis à cet effet et puisque seules deux espèces forment le genre. Une clé d’identification simplifiée et illustrée de la famille Ptychopteridae a été publiée par Fassbender (2014) dans le cadre de sa thèse de doctorat (2).
L’appartenance du spécimen à l’ordre des diptères (Diptera) est facilement établie par l’examen des ailes. Alors que la paire antérieure permet le vol, les ailes postérieures sont modifiées en haltères et servent des fonctions de balancier et de proprioception (Figure 1). L’aspect plutôt mince et allongé de l’insecte dans son ensemble (corps, pattes et antennes) permet de l’associer au sous-ordre des nématocères (Nematocera).

L’absence d’ocelles et la présence d’un lobe additionnel pré-haltère à la base de l’haltère (Figure 2) est indicateur de la famille Ptychopteridae. Cette structure n’est en effet retrouvée chez aucun autre diptère (3). On remarque également que le 2e segment abdominal est allongé (2).

La sous-famille Bittacomorphinae se caractérise par la présence de larges bandes blanches et noires sur les pattes, par l’absence de la première cellule médiane (M1) de l’aile (Figure 3) ainsi que par les antennes comprenant un minimum de 17 flagellomères (2).

On distingue le genre Bittacomorpha par le fort gonflement des basitarses des pattes (Figure 4). Les antennes portent moins de 20 flagellomères. Le genre ne comprend que deux espèces ; on distingue B. clavipes de B. occidentalis par la présence d’une bande grisâtre traversant le thorax longitudinalement en entier, se prolongeant après la suture thoracique transverse en «V» (Figure 5) (2).


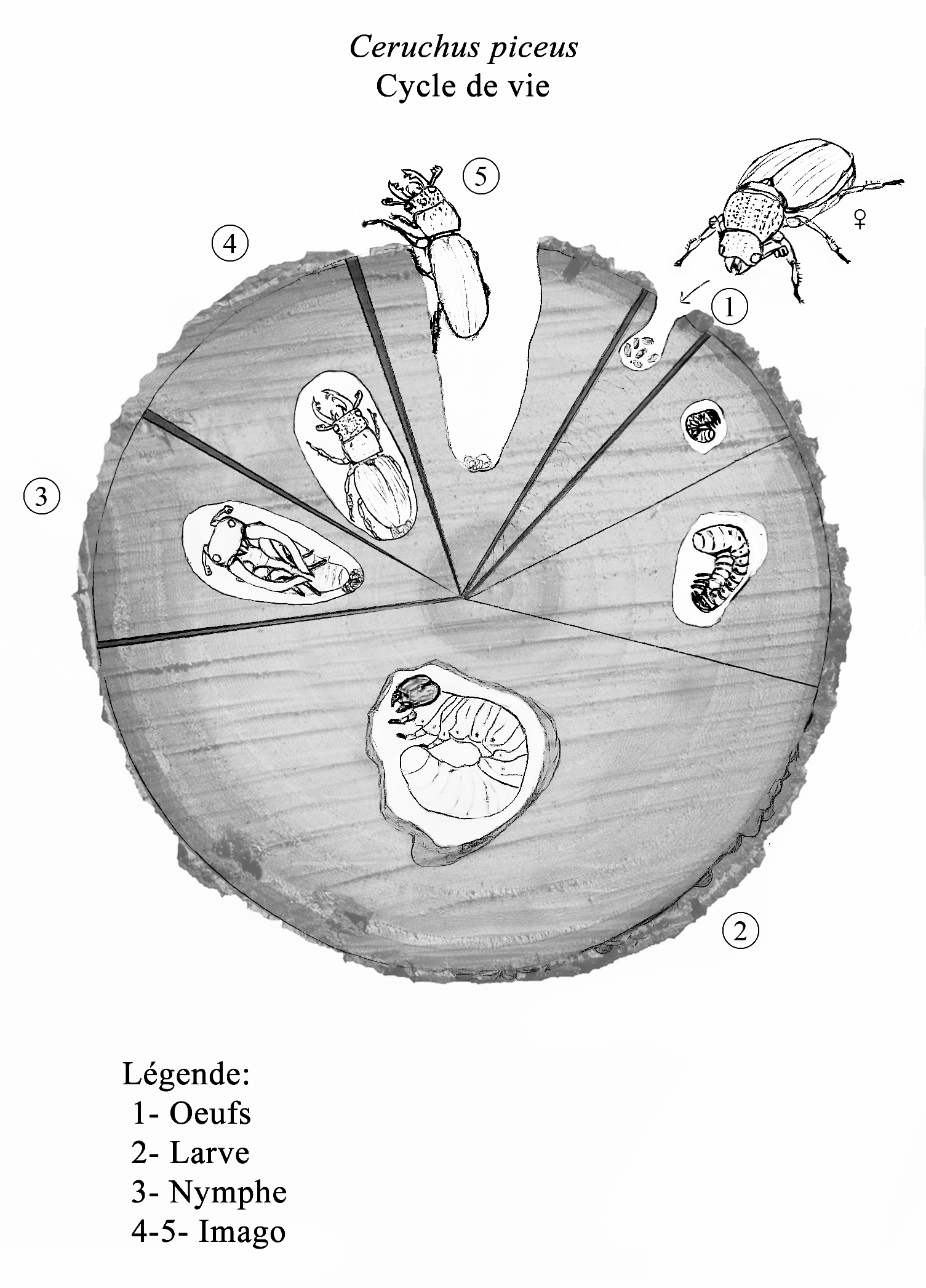
CYCLE DE VIE
La femelle B. clavipes dépose ses oeufs dans les sédiments humides de marais, marécages et fossés boueux qu’occuperont également la larve et la nymphe. Comme la respiration se fait toujours à l’air libre pendant cette période, les stades immatures ont développé un long siphon respiratoire, structure spécialisée en forme de tube comprenant une paire de spiracles terminaux et partiellement rétractile chez la larve. La présence de microstructures et de trois paires de pseudopodes abdominaux facilitent les mouvements de celle-ci dans les sédiment épais, notamment lors des mues qui ont lieu en surface pour ses 4 instars (Figure 6) (1,4). On remarque également que la tête de la larve est bien sclérifiée et que son corps allongé, originellement plutôt terne, peut prendre des colorations marquées suivant l’exposition aux minéraux présents dans le substrat (4). Son mode d’alimentation est filtreur de microparticules. B. clavipes passe la majorité de sa vie sous forme larvaire d’autant plus que l’hivernation a lieu lors du dernier instar (4) tandis que le stade nymphal ne dure que quelques semaines.

Le fantôme des marais adulte est une créature éphémère, d’une existence si courte et axée sur la reproduction que son alimentation est considérée facultative (4). Les pièces buccales sont réduites mais on peut observer de longs palpes maxillaires ainsi qu’un labre épais qui lui permettent de consommer des solutions sucrées telles du nectar ou du miellat (4). Dans certaines régions plus tempérées, il peut y avoir plusieurs générations de B. clavipes par année (i.e. espèce plurivoltine) (1).
ÉCOLOGIE
Le fantôme des marais est majoritairement retrouvé en habitats humides aux substrats riches en sédiments organiques et végétaux en décomposition tels que des marais ou marécages. Ils peuvent aussi se retrouver légèrement submergés sous la mince couche d’eau courante d’un ruisseau peu profond. On considère que son habitat optimal est un marécage au sol saturé et de substrat recouvert par un fin filet d’eau et riche en oxyde de fer (2).
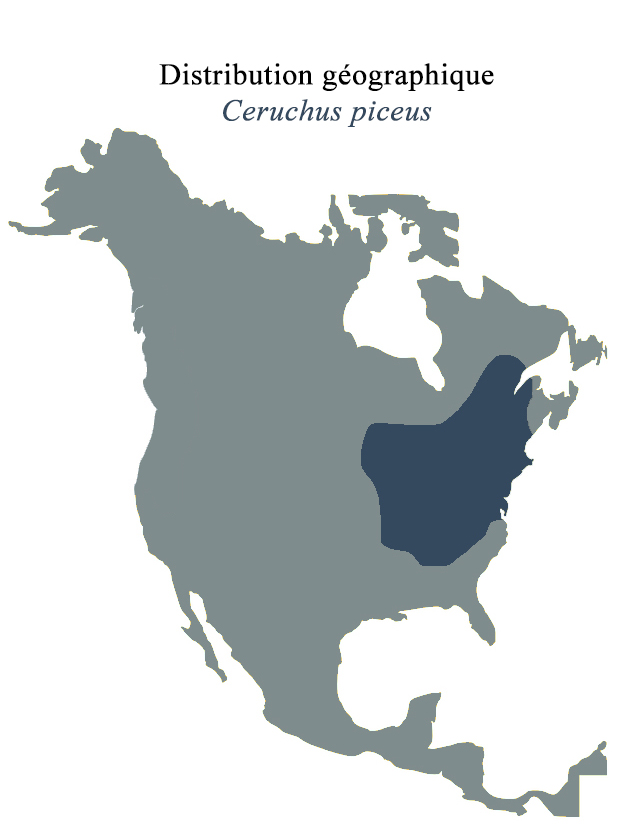
B. clavipes est l’espèce de Ptychopteridae dominante à travers toute l’Amérique du Nord et est principalement abondante dans l’est. Sa dispersion couvrant jusqu’à la moitié du continent peut s’expliquer par sa capacité à profiter des courants d’air pour traverses des barrières géologiques mineures grâce à son vol léger et flottant (2). Sa répartition à la frontière nord (région de Terre-neuve-et-Labrador) est limitée en raison des étés plus courts et principalement du manque de sédiments nécessaires au développement larvaire. La limite ouest (de l’est de la Colombie Britannique au nord est de l’Utah) est quant à elle plus difficile à déterminer; il n’y a pas encore consensus quant aux facteurs responsables bien que certains avancent à nouveau l’hypothèse du manque de sédiments appropriés (2). Bien que la limite sud du fantôme des marais soit établie comme au niveau du Texas, les données sont plutôt fragmentées et une population a même été répertoriée au Costa Rica, atteignant la zone néotropicale (2).
Références
- BOWLES, D.E. (1998). Life history of Bittacomorpha clavipes (Fabricius)(Diptera: Ptychopteridae) in an Ozark spring, USA. Aquatic Insects, 20(1), 29-34. http://www.tandfonline.com/doi/abs/10.1076/aqin.20.1.29.4486
- FASBENDER, A. (2014). Phylogeny and diversity of the phantom crane flies (Diptera: Ptychopteridae). Graduate Theses and Dissertations. Iowa State University, Paper 14133. http://lib.dr.iastate.edu/etd/14133/
- LUKASHEVICH, E.D. (2008). Ptychopteridae (Insecta: Diptera): History of its study and limits of the family. Paleontological Journal, 42(1), 66-74. http://link.springer.com/article/10.1007/s11492-008-1011-1
- ZWICK, P. (2004). Insecta: Diptera, Ptychopteridae. In Freshwater Invertebrates of the Malaysian Region (pp. 621-625). Aura Productions. https://www.researchgate.net/profile/Catherine_Yule/publication/233727128_44_Diptera_Ptychopteridae/links/09e4150acd05907295000000.pdf









































 Bibliographie
Bibliographie






{kind=link}
.jpg){kind=link}