
La guêpe à taches blanches
par Alys-Anh MORIN et Andrée-An THERRIEN
Texte et images ©2017 CC BY-SA 4.0, les auteurs

Classification
Classe: Insecta
Ordre: Hyménoptera
Sous-ordre: Apocrita
Infra-ordre: Aculeata
Super-famille: Vespoidea
Famille: Vespidae
Sous-famille: Vespinae
Genre: Dolichovespula
Espèce: Dolichovespula maculata (Linnaeus, 1763)
Identification du spécimen
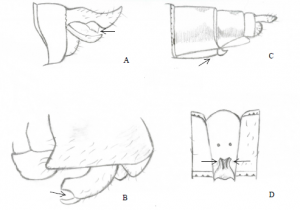
L’identification du spécimen consistait d’abord à observer ses caractéristiques particulières afin de trouver l’ordre d’insectes auquel il appartient. Le fait que le spécimen avait les ailes antérieures plus grandes que les ailes postérieures et qu’il y avait la présence de petits crochets sur ces dernières nous a permis de conclure qu’il appartient à l’ordre des Hyménoptères. Les ailes sont translucides et de teinte brunâtre. Les crochets servent à accrocher les ailes postérieures à celles antérieures afin de rendre le vol plus efficace. Le spécimen possède aussi trois ocelles derrière la tête pour capter la lumière et possiblement stabiliser le vol.



Ensuite, il a été possible de déterminer le sous-ordre Apocrita du spécimen grâce à la taille mince visible séparant le thorax du gastre. Cette taille permet de rendre le gastre plus flexible dans l’utilisation de l’oviscapte pour la ponte chez les «Parasitica» (guêpes parasitoïdes pondant leurs œufs à l’extérieur ou à l’intérieur d’un hôte où ceux-ci se développent) ou dans l’utilisation du dard comme mécanisme de défense chez l’infra-ordre Aculeata comprenant les guêpes piqueuses et sociales (Richter, 2000). En fait, les Aculeata auraient évolué d’un ancêtre parasitoïde généralisé par la modification de l’oviscapte (organe de ponte) en dard. En ce qui concerne le spécimen, il est possible de voir un dard ressorti et donc, il fait partie de l’infra-ordre Aculeata. Le spécimen appartient à la famille des Vespidae, car celle-ci comprend les guêpes sociales. Plus précisément, il fait partie de la sous-famille Vespinae puisqu’il est reconnu pour fabriquer des nids de carton ou de papier (Borror et White, 1970, p. 348 et 349).

Le genre Dolichovespula a pu être déterminé en observant l’espace présent entre la mandibule et l’œil du spécimen (espace oculo-malaire) (Species Dolichovespula maculata – Bald-faced Hornet, 2017). Les autres guêpes n’appartenant pas à ce genre ont normalement les mandibules accolées à leurs yeux, mais ce n’est pas le cas ici. Finalement, l’espèce du spécimen est D. maculata puisqu’il a le corps noir avec des lignes et bandes blanches ornant la tête, le thorax et les derniers segments de l’abdomen (Dubé, 2017). Justement, Dolichovespula maculata est communément appelée la « guêpe à taches blanches » à cause de cela.


Distribution géographique et habitat
Il existe 7 à 8 espèces appartenant au genre Dolichovespula en Amérique du Nord (Jacobs, 2010). Dolichovespula maculata est une espèce de guêpes native de l’Amérique du Nord, soit du Canada. Elle est retrouvée dans pratiquement toutes les provinces et territoires canadiens à l’exception du Nunavut (Buck et al., 2008). Elle habite aussi la côte occidentale et la plupart de la côte orientale des États-Unis. Toutefois, elle est surtout retrouvée au sud-est des États-Unis. Cette espèce préfère les zones tempérées naturelles, comme les forêts ou les roches, mais aussi les zones de végétation dans les endroits urbains afin de construire son nid. Par exemple, il est possible de retrouver un nid appartenant à cette espèce près d’une maison, d’une cabane ou même près de poteaux électriques (Buck et al., 2008). Son nid est construit dans un arbre, un arbuste ou tout autre endroit au-dessus du sol variant entre 1,1 et 20, 0 mètres de hauteur (Archer, 2006). Dolichovespula maculata est reconnue pour fabriquer son nid plus haut que la majorité des autres espèces de guêpes (Buck et al., 2008).
Biologie et cycle de vie
L’alimentation des guêpes à taches blanches varie selon le stade de développement de l’individu ainsi que l’endroit géographique où elles vivent. Les larves s’alimentent du nectar des fleurs rapporté et fourni par les adultes. Ces derniers sont principalement carnivores et se nourrissent de plusieurs types d’insectes et d’arthropodes vivants plus petits, telles que les araignées. Par contre, il arrive parfois qu’elles se nourrissent d’insectes plus gros comme des cigales ou des mantes prédatrices. De plus, elles peuvent occasionnellement fouiller de la viande morte pour des protéines, mais la majorité du temps, elles attrapent leurs proies vivantes. Pour discriminer les proies potentielles des proies non potentielles, elles survolent à basse altitude l’environnement et la végétation. Plus précisément, elles utilisent des repères visuels et se dirigent vers des silhouettes irrégulières lorsqu’elles en aperçoivent (Richter, 2000). Pour attraper leurs proies, les guêpes à taches blanches (et autres guêpes sociales) ont tendance à tuer leurs proies par une morsure plutôt que par leur dard. Celui-ci n’est seulement qu’utilisé comme moyen de défense en injectant un venin à l’intérieur de leur assaillant. Les guêpes à taches blanches se nourrissent aussi de fruits ou de certaines sèves d’arbres.

Le cycle de vie de la guêpe Dolichovespula maculata se base sur le cycle de vie de la colonie, se divisant en trois stades, et s’étale sur une année. À chaque printemps (début mai), les nouvelles colonies sont fondées par une reine née l’été précédent. La durée de vie d’une colonie, qui est d’environ quatre mois, s’étend jusqu’à la fin du mois de septembre. Par contre, à des latitudes plus basses, la longévité d’une colonie peut s’étendre jusqu’à 5 mois (Archer, 2006). Le premier stade de la colonie est sa fondation par une reine sortant d’hivernation. Cette dernière élève seule la première couvée d’ouvrières. Cette période dure entre 23-24 jours. Les œufs éclosent 6 jours après la ponte, puis les larves grandissent pendant 8 jours. Les ouvrières émergent une dizaine de jours plus tard. De façon surprenante, le couvain peut contribuer à la croissance de la colonie avant qu’il soit mature. En effet, il produit de la chaleur et fourni de la salive pour la reine. D’ailleurs, la construction du nid se fait à partir de bois mâché avec de l’amidon et de la salive. Cette préparation, qui est ensuite étendue autour de leurs pattes et leurs mandibules, sèche pour former du papier. Une fois les ouvrières adultes, le second stade débute. C’est la plus longue phase de vie de la colonie (Greene, 1984). Les ouvrières s’occupent de toutes les tâches d’entretien de la ruche (ex: construction d’alvéoles, collecte de nourriture et de matériaux pour la construction du nid) ainsi que les soins des futures ouvrières afin d’assurer la croissance de la colonie. De son côté, la reine se consacre entièrement à la reproduction en déposant des œufs dans les alvéoles de la ruche (Felippotti, 2009). Le dernier stade débute lorsqu’un certain seuil de croissance de la ruche est atteint. C’est à ce moment que la colonie concentre ses efforts sur la production d’individus reproducteurs tels que les reines et les mâles (Greene, 1984). La production de ces individus peut commencer durant le second stade, mais elle est très coûteuse. En effet, les mâles et les jeunes reines nourrissent uniquement les proies que les ouvrières rapportent dans la ruche. Conséquemment, les efforts déployés à la production de nouvelles ouvrières sont moindres.
La colonie est divisée en plusieurs castes : les ouvrières, le mâles et les reines. La différenciation est le résultat d’une nutrition particulière combinée à des alvéoles de plus grande taille. C’est pour cette raison que les reines D. maculata peuvent être de tailles différentes. Il peut arriver que certaines reines aient la même taille que des ouvrières d’une colonie voisine. Cependant, les reines (18-20 mm) sont toujours de plus grande taille que les ouvrières (12-14 mm) de leur propre colonie (Jacobs, 2010). La grande taille des reines favorise leur survie à l’hiver lorsqu’elles entrent en diapause. La diapause est un état dans lequel la guêpe diminue au minimum ses activités. Elle doit résister au gel tout en conservant suffisamment de réserves (Felippotti, 2009).
Bibliographie
ARCHER, Michael. (2006). Taxonomy, distribution and nesting biology of species of the genus Dolichovespula (Hymenoptera, Vespidae). Entomological Science, 9: 281–293
BORROR, Donald J. et WHITE, Richard E. (1970). Peterson Field Guides – Insects of America North of Mexico (1ère édition). Boston, États-Unis: Houghton Mifflin Company
BUCK, Matthias, MARSHALL, Stephen A. et CHEUNG, David K. B. (2008). Identification Atlas of the Vespidae (Hymenoptera, Aculeata) of the northeastern Neartic region. Repéré à https://cjai.biologicalsurvey.ca/bmc_05/84d_maculata.html
BUGGUIDE. Species Dolichovespula maculata – Bald-faced Hornet. (2017). Repéré à https://bugguide.net/node/view/2890
DUBÉ, Jocelyn. (2017). La guêpe à taches blanches – Dolichovespula maculata. Repéré à http://www.bestioles.ca/insectes/guepe-a-taches-blanches.html
ENCYCLOPEDIA OF LIFE. Dolichovespula maculata – Bald-faced Hornet. (s. d.). Repéré à http://eol.org/pages/239818/details
FELIPPOTTIA, G.T., TANAKA JUNIORA, G.M., NOLL, J.B. & WENZEL, J.W. (2009). Discrete dimorphism among castes of the bald-faced hornet Dolichovespula maculata (Hymenoptera: Vespidae) in different phases of the colony cycle. Journal of Natural History, 43(39-40): 2481-2490
GREENE, Albert. (1984). Production Schedules of Vespine Wasps: An Empirical Test of the Bang-Bang Optimization Model. Journal of The Kansas Entomological Society, 57(4):545-568
JACOBS, Steve. (2010). Insect Advice from Extension – Baldfaced Hornet. Repéré à http://ento.psu.edu/extension/factsheets/baldfaced-hornet
RICHTER, Raveret. (2000). Social Wasp (Hymenoptera: Vespidae) Foraging Behavior. Annual Review of Entomology, 45:121-150














































{kind=link}
{kind=link}
{kind=link}