Photo 1. Spécimen capturé grâce à un filet fauchoir le 7 septembre 2017 sur le chemin menant au lac Gaie à la Station de Biologie des Laurentides, Québec, Canada.
Ordre: Lépidoptera
Famille: Geometridae
Sous-famille: Ennominae
Genre:Lambdina
Espèce:Lambdina fiscellaria (Guenée, 1858) [1]
Identification:
Avec ses deux paires d’ailes recouvertes d’écailles on reconnaît instantanément que ce spécimen fait partie de l’ordre très diversifié des lépidoptères. De plus, sa couleur terne et ses antennes composées donnant l’aspect de plumes font penser qu’il figure parmi les papillons généralement nocturnes ou crépusculaires.
C’est grâce à sa couleur d’un beige pâle, au point noir présent sur chaque aile et surtout aux deux lignes transversales des ailes que l’identification à l’espèce Lambdina fiscellaria a été faite [4]. Effectivement, même si la couleur des individus de cette espèce peut varier du beige pâle au gris pâle et que ce ne sont pas tous les spécimens qui présentent le point noir sur les ailes postérieures, tous ont une ligne extérieure traversant les deux paires d’ailes et une ligne intérieure traversant seulement l’aile antérieure. De plus, tous ont un point noir sur chaque aile antérieure. Ces lignes et ces points sont également toujours symétriques sur les deux ailes [4].
Photo 2. Vu de côté des ailes du spécimen. À remarquer: les lignes transversales sur les ailes et le point des ailes antérieures.Photo 3: Vu de face du spécimen. À remarquer: la symétrie des lignes et des points sur les ailes ainsi que les antennes composées.
Le nom de la famille de Geometridea, dont il fait parti, est un dérivé de l’ancien grec où “Geo” signifie terre et “metridae” réfère à la mesure métrique, donnant ainsi la signification « mesure de la terre ». Cela fait référence au mouvement lobés des chenilles de cette famille, faisant ainsi penser que la chenille mesure son chemin [2]. Ces chenilles arpenteuses se déplacent ainsi puisqu’elles n’ont que 3 paires de fausses pattes abdominales [3].
Par sa couleur et sa forme, Besma quercivoraria est une espèce très ressemblante à Lambdina fiscellaria. Il serait même très facile de les confondre, les deux faisant partie de la même famille. Cependant, Besma quercivoraria se distingue par des lignes longitudinales sur les ailes leurs donnant un aspect de « coquillage ». Également, Besma quercivoraria vole au début de l’été, tandis que Lambdina fiscellaria est observé en automne [5].
Photo 6 : Besma quercivoraria vu de dessus. À remarquer: les lignes longitudinales sur toutes les ailes. Photo par Andy Reago et Chrissy McClarren originalement publiée sur le site de Flickr, selon le code légal de Creative commons.Photo 7: Photo de Lambdina fiscellaria. À remarquer: Absence de lignes longitudinales (contrairement à Besma quercivoraria)
Répartition géographique:
Lambdina fiscellaria est une espèce indigène de l’Amérique du Nord [6]. On le retrouve dans toutes les provinces du Canada ainsi qu’au Nord des états Unis [7]. Au Québec, il est présent dans toutes les régions, de préférence dans les boisés humides et frais de conifères, mais également dans les boisés mixtes et les érablières [4].
Cette espèce n’est associé à aucun statut de précarité [8]. Elle est en fait très présente dans les forêts Canadiennes et est considérée comme un des plus importants défoliateurs de conifères du continent. Les épidémies apparaissent et disparaissent sporadiquement sur tout le territoire englobant le Canada et le Nord des États-Unis [9].
Alimentation:
Les chenilles se nourrissent entre autres des feuilles d’arbustes et d’arbres, comme le bouleau et l’érables, ainsi que des aiguilles de plusieurs conifères, comme l’épinette blanche ou le mélèze laricin. Même si la chenille préfère le sapin baumier ainsi que la pruche du Canada, d’où son appellation « Arpenteuse de la pruche », elle peut se nourrir d’autres arbres, particulièrement lors de période épidémique[6].
Cycle de vie:
Pour l’Arpenteuse de la Pruche, la ponte a lieu entre la fin août et octobre, préférablement sur la mousse et le lichen qui recouvrent les arbres. Souvent, les œufs sont regroupés par deux ou trois. L’éclosion des œufs se produit au printemps, suite à l’éclosion des bourgeons de sapins. Les chenilles de Lambdina fiscellaria font du phototropisme positif, c’est-à-dire qu’elles vont être attirées par la lumière. Elles vont donc se diriger vers l’extrémité des branches ainsi qu’à la cime des arbres et vont commencer à se nourrir des épines de leur hôte. Tout au long du développement de la chenille, soit lors de ses différents stades larvaires, ses préférences alimentaires vont changer. Commençant par les bourgeons fraîchement éclos, puis se nourrissant des feuillages plus vieux arrivé aux derniers stades larvaires. Les chenilles mangent rarement une aiguille complètement, la laissant rougir, et montrant ainsi les traces de son passage. Suite à son développement larvaire, la formation de la chrysalide commence. L’adulte en émerge par la suite et sera présent de la fin août jusqu’à tardivement à l’automne. Inactif le jour, ce n’est que le soir venu que la recherche de partenaire s’entame. Une fois que le mâle trouve une femelle, le cycle recommence [10].
Impact environnemental
Comme mentionné plus tôt, Lambdina fiscellaria possède un fort caractère épidémique. Les infestations sont visibles grâce au rougissement des aiguilles des arbres atteints. Cela a pour conséquences de rendre les arbres les plus attaqués très vulnérables à d’autres facteurs environnementaux, comme les maladies, ce qui peut au final causer la mort des arbres.
Les infestations au Québec s’étendent habituellement sur une durée de 2 à 3 ans et sont espacées de 5 à 10 ans. La deuxième année est celle où la superficie affectée est la plus grande, mais il est rare que l’épidemie continue plus de 3 ans, car plusieurs variables tendent à la freiner naturellement. En effet, le manque de nourriture, les variations climatiques, la prédation, les maladies causées par des champignons et, principalement, l’attaque de guêpes parasitoïdes sur les oeufs au printemps permettent de diminuer les populations de chenilles et ainsi de réprimer les invasions de Lambdina fiscellaria [6][10].
[2] Robin McLeod, John; Jane Balaban, Beatriz Moisset & Chuck Entz (2009). BugGuide, Family Geometridae – Geometrid Moths. Consulté le 25 octobre 2017. URL:https://bugguide.net/node/view/188
Calligrapha philadelphica ou le Calligraphe ligné du cornouiller a été décrit par Linnaeus en 1758. Ce petit coléoptère fait partie de la grande famille des Chrysomelidae ou « Leaf beetles » qui compte plus de 35 000 espèces à travers le monde (Jolivet, 1988) et pourrait compter jusqu’à 60 000 espèces selon des estimations plus récentes (Reid, 1995) (Arnett, 2002). De ces nombreuses espèces, 14 sous-familles en sont les représentantes. Les chrysomèles sont herbivores et ont très souvent une plante hôte propre à l’espèce pour l’alimentation et la reproduction. Étant donné leurs modes d’alimentation phytophages, ils sont donc nuisibles pour les plantes dont ils se nourrissent. Les Chrysomelidae montrent une réduction ou perte totale de certaines structures en comparaison avec les autres familles. Par exemple, les structures buccales sont plus réduites car les premiers chrysomèles consommait du pollen, la transition de pollinivore (se nourrit de pollen) à phyllophage (se nourrit de feuilles) nécessite une complexité moindre (voir figure 2). Par ailleurs, plusieurs chrysomèles ont moins de veinures sur les ailes, l’hypothèse est que l’ancêtre aurait une taille beaucoup plus réduite (Reid, 1995).
Figure 2. Pièces buccales réduites du Calligrapha philadelphica
IDENTIFICATION et MORPHOLOGIE
Le genre Calligrapha est facilement identifiable grâce aux motifs caractéristiques sur les élytres (Zurita et al., 2004). Le patron générale des motifs, la couleur des élytres et du pronotum sont différents selon l’espèce. Les espèces C. vivina, C. suturella, ou C. multipunctata ressemble beaucoup à C. philadelphica sauf pour quelques détails.
Calligrapha philadelphica a une petite taille de 7 à 9 millimètres. Ils ont un pronotum entièrement noir et des pattes dotées de deux griffes bien définies (Majka, s.d.)(Arnett, 2002). Cette espèce ne présente aucune différence entre les mâles et les femelles, c’est-à-dire qu’il n’y a aucun dimorphisme sexuel.
Bien sûr, si on trouve le spécimen sur sa plante hôte (ici le cornouiller), cela aide à l’identification de l’espèce. Cependant, nous avons trouvé notre spécimen dans une Sarracenia (plante carnivore) dans la tourbière du Lac Geai. Nous avons donc identifier notre espèce grâce à une clef d’identification des calligraphes de l’est du Canada (14 espèces sur 38 en Amérique du Nord) (Majka, s.d.).
Les motifs noirs des élytres varient aussi entre individus et entre localités, c’est-à-dire qu’il y a beaucoup de variation intra- et inter-populations. Quand nous avons voulu confirmer l’identification de notre spécimen, nous l’avons comparé aux spécimens de la collection Ouellet-Robert, dont les motifs étaient légèrement différents prouvant la grande variation génétique.
C’est surtout la taille et l’épaisseur des motifs qui varient beaucoup, le calus huméral est plus fin que ceux des spécimen de la collection (Zurita et al., 2004). De plus, si on regarde le calus huméral, il peut parfois être fusionné avec la lunule humérale (en forme de lune) (Brown, 1945). Sur notre spécimen, on retrouve les deux formes. En effet, ils sont fusionnés sur l’élytre gauche et séparé sur l’élytre gauche. La ligne parallèle à la suture des élytres varie aussi en longueur. Le point enfermé par la lunule peut être divisé en 2 points, ce qui est le cas chez notre spécimen (voir figure 3).
Figure 3. Caractéristiques principales sur les élytres du Calligrapha philadelphica
Afin de s’assurer de l’identification de l’espèce, il faut vérifier s’il y a des plantes du genre Cornus à l’endroit où nous l’avons trouvé. L’étude de Racine, en 2013, démontre qu’il y a présence de Cornus canadensis autour du lac Geai de la station de biologie des Laurentides.
BIOLOGIE
Les différentes espèces de calligraphes restreignent leur alimentation à quelques espèces de plantes (uniquement des dicotylédones). Calligrapha philadelphica est associé au genre Cornus, d’où son nom: calligraphe ligné du cornouiller. Le cornouiller est la plante hôte de cette espèce qui va donc passer la grande majorité de sa vie sur cette plante (alimentation, reproduction, développement des larves) (Robertson, 1966).
Figure 4. Cycle de vie de Calligrapha philadelphica. Dessin par Victoria Thelamon.
Dans l’Est du Canada, toutes les espèces de calligraphes semble avoir le même cycle de vie (voir figure 4). Les adultes hibernent et réapparaissent sur leur plante hôte dès le mois de Mai. La ponte débute vers la mi-Mai jusqu’au mois de juin. Les œufs sont jaune pâle, rose ou corail, déposés individuellement ou en amas, pouvant atteindre un total de 32 oeufs, sur les feuilles de la plante hôte (Robertson, 1966)(Brown, 1945). Les larves émergent et se nourrissent jusqu’à atteindre le dernier stade larvaire. Elles s’enfouissent ensuite dans le sol pour la pupation qui dure en moyenne 8 jours. Lorsque l’adulte émerge il est jaune pâle sans aucun motif sur ses élytres. Les couleurs apparaîtront peu à peu, devenant de plus en plus foncées et nettes avec le temps. Les premiers adultes de la nouvelle génération émergent de fin juillet jusqu’au mois de Septembre, ils chercheront ensuite un lieu pour l’hibernation. Une seule génération est produite par année (Brown, 1945).
Les populations de calligraphes sont souvent des colonies isolées, restreintes à la distribution de leur plante hôte. Les groupes peuvent être petits ou grand, et il peut éventuellement avoir des échanges (migration) entre les populations. Ainsi, il y a une grande variabilité entre les populations. Certaines espèces de Calligraphes ont même développé la parthénogenèse (C. philadelphica n’est pas concerné) (Brown, 1945).
DISTRIBUTION
Figure 5. Aire de répartition de Calligrapha philadelphica. Carte réalisée par Victoria Thelamon.
La biogéographie du Calligraphe ligné du cornouiller, dans la zone Néarctique dont il est endémique, suit presque identiquement la distribution de sa plante hôte (Cornus). Il préfère les boisés et les lisières qui longent les forêts des régions tempérées où se trouve le Cornus. Les individus ne s’aventurent jamais très loin de leur plante hôte. Au Canada, ses habitats de prédilection se retrouvent surtout dans les provinces de l’Est, mais on les retrouve également jusqu’en Colombie Britannique (voir figure 5). Aux États-Unis, on observe la même tendance soit que le Calligraphe ligné du cornouiller se retrouve majoritairement dans les états de la côte Est (du Maine jusqu’à la Géorgie), mais aussi dans quelques états de l’ouest comme Washington, l’Idaho et le Montana (Gómez-Zurita, 2005).
RÉFÉRENCES
Arnett, R. H., Thomas, M. C., Skelley, P. E., & Frank, J. H. (Eds.). (2002). American Beetles, Volume II: Polyphaga: Scarabaeoidea through Curculionoidea (Vol. 2). CRC press.
Brown, W. J. (1945). Food-plants and distribution of the species of Calligrapha in Canada, with descriptions of new species (Coleoptera, Chrysomelidae). The Canadian Entomologist, 77(7), 117-133.
Gómez-Zurita, J., Vogler, A. P., & Funk, D. J. (2004). Diagnosing an overlooked North American taxon: Biological observations and mitochondrial insights on Calligrapha suturella Schaeffer, new status (Coleoptera, Chrysomelidae). Annals of the Entomological Society of America, 97(1), 28-36.
Gómez-Zurita, J. (2005). New Distribution Records and Biogeography of Calligrapha Species (Leaf Beetles), in North America (Coleoptera: Chrysomelidae, Chrysomelinae). The Canadian Field-Naturalist, 119(1), 88-100.
Jolivet, P., Petitpierre, E., Hsiao, T.H. (1988). Biology of Chrysomelidae (vol 42). Kluwer Academic Publishers.
Reid, C. A. M. (1995). A cladistic analysis of subfamilial relationships in the Chrysomelidae sensu lato (Chrysomeloidea). Biology, phylogeny and classification of Coleoptera: papers celebrating the 80th birthday of Roy A. Crowson, 2, 559-631.
Robertson, J. G. (1966). The chromosomes of bisexual and parthenogenetic species of Calligrapha (Coleoptera: Chrysomelidae) with notes on sex ratio, abundance and egg number. Canadian Journal of Genetics and Cytology, 8(4), 695-732.
Figure 1. Vue dorsale du stade adulte de l’espèce Platydracus violaceus
Spécimen attrapé à la Station de Biologie des Laurentides en date du 7 septembre 2017. Le spécimen, prélevé de jour en milieu humide, reposait inactif sous une souche d’arbre en décomposition. Lors de son échantillonnage à la main, le spécimen exposait un comportement dissuasif en vibrant et en remuant l’abdomen comme pour piquer.
Classification
Selon la clé de l’article Staphylinidae of Eastern Canada and Adjacent United States (Brunke et al. 2011)
L’ordre des coléoptères, représentant 40 % de toutes les espèces d’arthropodes décrites et dont le nombre d’espèces existant est estimé à 1,5 million, regroupe de nombreuses familles, dont celle des Staphylinidae (Stork et al., 2015). Comptant parmi plus de 55 440 espèces décrites et regroupées dans plus de 3200 genres, la famille des Staphylinidae constitue l’une des plus grandes familles de coléoptères (Grebennikov et al., 2009). Alors que l’histoire fossile de ce groupe remonte à quelque 200 millions d’années, ce sont, à ce jour, près de 400 nouvelles espèces qui sont encore mondialement répertoriées par an. En Amérique du Nord, la famille des Staphylinidae, regroupant près de 4100 espèces à travers 523 genres, forme la plus grande famille de coléoptère présente en ces territoires (Arnett et al., 2001).
Biologie de l’insecte
Morphologie générale de la famille des Staphylinidae
La famille des Staphylinidae se distingue facilement des autres familles de coléoptères par leur corps allongé, linéaire et souple ainsi qu’à leurs élytres, très raccourcis, laissant à découvert la plus grande partie de l’abdomen : de 3 à 6 segments abdominaux sont laissés à découvert. Sous ces élytres se trouvent des ailes bien développées conférant à cette famille une bonne capacité de vol (Chagnon et al., 1962). Bien que la famille soit confondue avec l’ordre des dermaptères, les Staphylinidae ne portent pas de pinces à la pointe de leur abdomen et possèdent des antennes de 11 segments ou moins (Klimaszewski et al. 1996). Leurs petits corps, mince et étroit, ainsi que leur abdomen, flexible, permet aux Staphylinidae de coloniser des habitats telle la litière, inaccessible à des organismes plus robustes et moins flexibles. Bien que certaines espèces possèdent des couleurs vives aux reflets métalliques, la majorité des Staphylinidae, dont la taille varie entre 0,7 mm et 25 mm, présentent des couleurs plutôt sombres (marron et noir) se fondant à celles de la canopée (White 1983). La plupart des espèces, lorsqu’inquiétées, adoptent un comportement dissuasif en relevant et remuant leur abdomen comme pour piquer : ces espèces sont pourtant inoffensives, bien que certains sous-groupes de la famille possèdent des glandes abdominales et des produits chimiques défensifs (Cline 2011).
Écologie
Aussi appelés « Rove Beetles« , les Staphylinidae sont parmi les coléoptères les plus performants quant à leur capacité à coloniser une grande variété d’habitats. Quoique pauvrement représentés en milieux aquatiques, ceux-ci sont communs dans tous les biomes. Alors que certaines espèces se retrouvent en milieux forestier, d’autres encore habitent les forêts tropicales, la toundra arctique ainsi que les régions alpines et les habitats océaniques intertidal et sublittoral tels les tourbières, les marais et divers autres milieux humides (Campbell et al. 1991). Or, c’est environ la moitié de toutes les espèces de Staphylinidae qui vivent en milieux forestiers. Celles-ci possèdent un large éventail d’habitats allant du long des rivages à la litière du sol, du dessous de l’écorce des arbres aux matières stercoraires et des cadavres. D’autres encore se retrouvent sur les champignons et les fleurs ; certaines espèces, myrmécophiles ou termitophiles, vont même jusqu’à habiter les fourmilières et les termitières de fourmis et de termites. Le cycle de vie de ces espèces, dont la plupart surviennent dans le sol, fait de cette famille l’une des plus importantes de la pédofaune (Newton et al. 1990). La plupart des espèces contenues dans la famille des Staphylinidae évitent tous contacts directs avec la lumière et sont généralement nocturnes, ce mode de vie offrant une protection plus grande contre la prédation et la dessiccation (Klimaszewski et al. 1996).
Ils sont d’agiles prédateurs généralistes se nourrissant d’une grande variété d’arthropodes et d’invertébrés terrestres. Certaines espèces sont cependant spécialisées pour s’attaquer spécifiquement à certains groupes d’insecte. D’autres encore sont saprophages et ont pour ressources alimentaires diverses matières en décomposition ; algues, pollen et inflorescences florales. D’autre encore sont phytophages et se nourrissent d’organismes vivants, tels les champignons, desquels ils se nourrissent des hyphes et des spores (Cline 2011).
Figure 2.Vue dorsale des setae, poils qui ont pour fonction de faciliter le déplacement de l’espèce grâce à une meilleure adhérence au substrat.
Où retrouve-t-on l’espèce Platydracus violaceus ?
Platydracus violaceus est une espèce habitant les milieux forestiers. Elles habitent autant les milieux humides qu’offre la canopée et la litière, autant ceux offerts par le dessous de l’écorce de feuillus morts, tels les chênes, les marronniers et les érables. Quoiqu’il soit aussi possible de les retrouver sous l’écorce des pins blancs ainsi que dans le bois pourri, dans certains champignons ou carcasses en décomposition ainsi que sur les excréments, ces cas sont moins fréquents et ne constituent pas les habitats préférentiels de l’espèce (Brunke et al. 2011). L’hiver, ces espèces hibernent pour survivre aux températures plus froides. Les larves, qui éclosent au printemps, partagent le même habitat que les adultes et se nourrissent de diverses larves pontent par d’autres insectes (Arnett et al. 2001).
Au Canada, alors qu’encore bien des espèces demeurent encore inconnues, ce sont 1 048 espèces de la famille des Staphylinidae qui ont été répertoriées, lesquels peuvent occuper un large territoire tel le montre la distribution de l’espèce Platydracus violaceus (Campbell et al. 1991).
L’identification du spécimen a été effectuée selon la clé de l’article Staphylinidae of Eastern Canada and Adjacent United States (Brunke et al. 2011). L’identification requiert l’étude de traits subtils ayant permis la distinction de l’espèce parmi la famille des Staphylinidae.
Les figures 4 et 5, annotées, relaient les divers traits caractéristiques ayant permit l’identification du spécimen Platydracus violaceus.
Tout d’abord, l’identification de la sous-famille Staphylininae a été permise grâce à l’absence de coxa postérieur (a).
Ensuite, la combinaison des quelques caractéristiques suivantes a permis de mettre en évidence la tribu Staphilini : les élytres se rencontrant à la ligne médiane du corps (b), un cou large mesurant la moitié de la tête (c) et la base des antennes séparées par une distance plus grande que celle séparant les yeux de la base des antennes (d).
Notre spécimen possédait cinq tarsomères par tarse (e), des palpes maxillaires et labiaux courts (f), des ponctuations sur la surface dorsale du cou (g) et une tête au bout étroit plus large que la moitié de la longueur du pronotum (h). Ces caractéristiques nous ont permis d’identifier la sous-tribu Staphilinina.
Finalement, notre spécimen fait partie du genre Platydracus, car il possède un pronotum ponctué aux angles ronds (i) avec une ligne non ponctuée médiane en son centre (j). De plus, les setae (poils) situés sur les côtés de la tête ne sont pas limités à la moitié antérieure de la tempe (k).
Les caractéristiques les plus remarquables de Platydracus violaceus sont les élytres violets aux reflets métalliques et les poils pâles recouvrant certaines parties de l’abdomen. Il est possible de confondre Platydracus violaceus avec Platydracus viridanus, ce dernier ayant des élytres violets lorsqu’il est exposé à des substances toxiques se trouvant dans des trappes à insectes. Dans le cas de notre spécimen, capturé dans son habitat naturel, aucune exposition à un agent n’a eu lieu. De plus, P. violaceus possède des pattes entièrement noires et un pronotum aussi large que la tête, ce qui n’est pas le cas de P. viridanus (Brunke et al. 2011).
Figure 6. Vue dorsale de l’abdomen de l’espèce Platydracus violaceus ainsi que des poils caractéristiques les recouvrant.
Références:
Arnett Jr, R. H., & Thomas, M. C. (Eds.). (2000). American Beetles, Volume I: Archostemata, Myxophaga, Adephaga, Polyphaga: Staphyliniformia. CRC Press, Boca Raton, FL.
Brunke A., Newton A., Klimaszewski J., Majka C. Et Marshall S. (2011). Staphylinidae of Eastern Canada and Adjacent United States. Key to Subfamilies ; Staphylininae : Tribes and Subtribes, and Species of Staphylinina. Canadian Journal of Arthropod Identification 12., pp.1-111. DOI: 10.3752/cjai.2011.1
Chagnon, G. et Robert, A. (1962) .Principaux coléoptères de la province de Québec (2e éd.). Montréal, Canada : Les Presses de l’Université de Montréa
Grebennikov, V. V., & Newton, A. F. (2009). Good-bye Scydmaenidae, or why the ant-like stone beetles should become megadiverse Staphylinidae sensu latissimo (Coleoptera). European Journal of Entomology, 106(2), 275-301
Klimaszewski, J., Newton JR, A. F., & Thayer, M. K. (1996). A review of the New Zealand rove beetles (Coleoptera: Staphylinidae). New Zealand journal of zoology, 23(2), 143-160. DOI: 10.1080/03014223.1996.9518074
Levesque, C., Levesque, G.-Y. (1995). Abundance, Diversity and Dispersal Power of Rove Beetles (Coleoptera : Staphylinidae) in a Raspberry Plantation and Adjacent Sites in Eastern Canada. Journal of the Kansas entomological society, 68(3), 355-370
Cline, A. R. (2011). Checklist of the Beetles of Maine, USA. The Coleopterists Bulletin, 65(4), 364-365.
Newton, A. F., Jr. (1990). Insecta: Coleoptera Staphylinidae adults and larvae. In D. L. Dindal (ed.), Soil Biology Guide. Chap. 38, pp. 1137-1174. John Wiley and Sons, Inc., New York
Paulian, R. (1988). Biology of the Coleoptera. Editions Lechevalier
Stork, N. E., McBroom, J., Gely, C., & Hamilton, A. J. (2015). New approaches narrow global species estimates for beetles, insects, and terrestrial arthropods. Proceedings of the National Academy of Sciences, 112(24), 7519-7523.
White, R. E. (1998). The Beetles of North America (Vol. 29). Houghton Mifflin Harcourt.
L’identification du spécimen consistait d’abord à observer ses caractéristiques particulières afin de trouver l’ordre d’insectes auquel il appartient. Le fait que le spécimen avait les ailes antérieures plus grandes que les ailes postérieures et qu’il y avait la présence de petits crochets sur ces dernières nous a permis de conclure qu’il appartient à l’ordre des Hyménoptères. Les ailes sont translucides et de teinte brunâtre. Les crochets servent à accrocher les ailes postérieures à celles antérieures afin de rendre le vol plus efficace. Le spécimen possède aussi trois ocelles derrière la tête pour capter la lumière et possiblement stabiliser le vol.
Figure 1: Les ailes antérieures sont plus grandes que les ailes postérieures Figure 2: Les ailes antérieures sont accrochées aux ailes postérieures à l’aide de petits crochets Figure 3: Les trois ocelles situés derrière la tête de la guêpe
Ensuite, il a été possible de déterminer le sous-ordre Apocrita du spécimen grâce à la taille mince visible séparant le thorax du gastre. Cette taille permet de rendre le gastre plus flexible dans l’utilisation de l’oviscapte pour la ponte chez les «Parasitica» (guêpes parasitoïdes pondant leurs œufs à l’extérieur ou à l’intérieur d’un hôte où ceux-ci se développent) ou dans l’utilisation du dard comme mécanisme de défense chez l’infra-ordre Aculeata comprenant les guêpes piqueuses et sociales (Richter, 2000). En fait, les Aculeata auraient évolué d’un ancêtre parasitoïde généralisé par la modification de l’oviscapte (organe de ponte) en dard. En ce qui concerne le spécimen, il est possible de voir un dard ressorti et donc, il fait partie de l’infra-ordre Aculeata. Le spécimen appartient à la famille des Vespidae, car celle-ci comprend les guêpes sociales. Plus précisément, il fait partie de la sous-famille Vespinae puisqu’il est reconnu pour fabriquer des nids de carton ou de papier (Borror et White, 1970, p. 348 et 349).
Figure 4: La taille mince séparant le thorax du gastre chez les Apocrita. Le dard comme mécanisme de défense chez les Aculeata
Le genre Dolichovespula a pu être déterminé en observant l’espace présent entre la mandibule et l’œil du spécimen (espace oculo-malaire) (Species Dolichovespula maculata – Bald-faced Hornet, 2017). Les autres guêpes n’appartenant pas à ce genre ont normalement les mandibules accolées à leurs yeux, mais ce n’est pas le cas ici. Finalement, l’espèce du spécimen est D. maculata puisqu’il a le corps noir avec des lignes et bandes blanches ornant la tête, le thorax et les derniers segments de l’abdomen (Dubé, 2017). Justement, Dolichovespula maculata est communément appelée la « guêpe à taches blanches » à cause de cela.
Figure 5: L’espace oculo-malaire entre la mandibule et l’œil de la guêpe Figure 6: Les diverses taches blanches sur la tête, le thorax et l’abdomen de la guêpe
Distribution géographique et habitat
Il existe 7 à 8 espèces appartenant au genre Dolichovespula en Amérique du Nord (Jacobs, 2010). Dolichovespula maculata est une espèce de guêpes native de l’Amérique du Nord, soit du Canada. Elle est retrouvée dans pratiquement toutes les provinces et territoires canadiens à l’exception du Nunavut (Buck et al., 2008). Elle habite aussi la côte occidentale et la plupart de la côte orientale des États-Unis. Toutefois, elle est surtout retrouvée au sud-est des États-Unis. Cette espèce préfère les zones tempérées naturelles, comme les forêts ou les roches, mais aussi les zones de végétation dans les endroits urbains afin de construire son nid. Par exemple, il est possible de retrouver un nid appartenant à cette espèce près d’une maison, d’une cabane ou même près de poteaux électriques (Buck et al., 2008). Son nid est construit dans un arbre, un arbuste ou tout autre endroit au-dessus du sol variant entre 1,1 et 20, 0 mètres de hauteur (Archer, 2006). Dolichovespula maculata est reconnue pour fabriquer son nid plus haut que la majorité des autres espèces de guêpes (Buck et al., 2008).
Biologie et cycle de vie
L’alimentation des guêpes à taches blanches varie selon le stade de développement de l’individu ainsi que l’endroit géographique où elles vivent. Les larves s’alimentent du nectar des fleurs rapporté et fourni par les adultes. Ces derniers sont principalement carnivores et se nourrissent de plusieurs types d’insectes et d’arthropodes vivants plus petits, telles que les araignées. Par contre, il arrive parfois qu’elles se nourrissent d’insectes plus gros comme des cigales ou des mantes prédatrices. De plus, elles peuvent occasionnellement fouiller de la viande morte pour des protéines, mais la majorité du temps, elles attrapent leurs proies vivantes. Pour discriminer les proies potentielles des proies non potentielles, elles survolent à basse altitude l’environnement et la végétation. Plus précisément, elles utilisent des repères visuels et se dirigent vers des silhouettes irrégulières lorsqu’elles en aperçoivent (Richter, 2000). Pour attraper leurs proies, les guêpes à taches blanches (et autres guêpes sociales) ont tendance à tuer leurs proies par une morsure plutôt que par leur dard. Celui-ci n’est seulement qu’utilisé comme moyen de défense en injectant un venin à l’intérieur de leur assaillant. Les guêpes à taches blanches se nourrissent aussi de fruits ou de certaines sèves d’arbres.
Figure 7: Dolichovespula maculata en train de manger un arthropode
Le cycle de vie de la guêpe Dolichovespula maculata se base sur le cycle de vie de la colonie, se divisant en trois stades, et s’étale sur une année. À chaque printemps (début mai), les nouvelles colonies sont fondées par une reine née l’été précédent. La durée de vie d’une colonie, qui est d’environ quatre mois, s’étend jusqu’à la fin du mois de septembre. Par contre, à des latitudes plus basses, la longévité d’une colonie peut s’étendre jusqu’à 5 mois (Archer, 2006). Le premier stade de la colonie est sa fondation par une reine sortant d’hivernation. Cette dernière élève seule la première couvée d’ouvrières. Cette période dure entre 23-24 jours. Les œufs éclosent 6 jours après la ponte, puis les larves grandissent pendant 8 jours. Les ouvrières émergent une dizaine de jours plus tard. De façon surprenante, le couvain peut contribuer à la croissance de la colonie avant qu’il soit mature. En effet, il produit de la chaleur et fourni de la salive pour la reine. D’ailleurs, la construction du nid se fait à partir de bois mâché avec de l’amidon et de la salive. Cette préparation, qui est ensuite étendue autour de leurs pattes et leurs mandibules, sèche pour former du papier. Une fois les ouvrières adultes, le second stade débute. C’est la plus longue phase de vie de la colonie (Greene, 1984). Les ouvrières s’occupent de toutes les tâches d’entretien de la ruche (ex: construction d’alvéoles, collecte de nourriture et de matériaux pour la construction du nid) ainsi que les soins des futures ouvrières afin d’assurer la croissance de la colonie. De son côté, la reine se consacre entièrement à la reproduction en déposant des œufs dans les alvéoles de la ruche (Felippotti, 2009). Le dernier stade débute lorsqu’un certain seuil de croissance de la ruche est atteint. C’est à ce moment que la colonie concentre ses efforts sur la production d’individus reproducteurs tels que les reines et les mâles (Greene, 1984). La production de ces individus peut commencer durant le second stade, mais elle est très coûteuse. En effet, les mâles et les jeunes reines nourrissent uniquement les proies que les ouvrières rapportent dans la ruche. Conséquemment, les efforts déployés à la production de nouvelles ouvrières sont moindres.
La colonie est divisée en plusieurs castes : les ouvrières, le mâles et les reines. La différenciation est le résultat d’une nutrition particulière combinée à des alvéoles de plus grande taille. C’est pour cette raison que les reines D. maculata peuvent être de tailles différentes. Il peut arriver que certaines reines aient la même taille que des ouvrières d’une colonie voisine. Cependant, les reines (18-20 mm) sont toujours de plus grande taille que les ouvrières (12-14 mm) de leur propre colonie (Jacobs, 2010). La grande taille des reines favorise leur survie à l’hiver lorsqu’elles entrent en diapause. La diapause est un état dans lequel la guêpe diminue au minimum ses activités. Elle doit résister au gel tout en conservant suffisamment de réserves (Felippotti, 2009).
Bibliographie
ARCHER, Michael. (2006). Taxonomy, distribution and nesting biology of species of the genus Dolichovespula (Hymenoptera, Vespidae). Entomological Science, 9: 281–293
BORROR, Donald J. et WHITE, Richard E. (1970). Peterson Field Guides – Insects of America North of Mexico (1ère édition). Boston, États-Unis: Houghton Mifflin Company
BUCK, Matthias, MARSHALL, Stephen A. et CHEUNG, David K. B. (2008). Identification Atlas of the Vespidae (Hymenoptera, Aculeata) of the northeastern Neartic region. Repéré à https://cjai.biologicalsurvey.ca/bmc_05/84d_maculata.html
FELIPPOTTIA, G.T., TANAKA JUNIORA, G.M., NOLL, J.B. & WENZEL, J.W. (2009). Discrete dimorphism among castes of the bald-faced hornet Dolichovespula maculata (Hymenoptera: Vespidae) in different phases of the colony cycle. Journal of Natural History, 43(39-40): 2481-2490
GREENE, Albert. (1984). Production Schedules of Vespine Wasps: An Empirical Test of the Bang-Bang Optimization Model. Journal of The Kansas Entomological Society, 57(4):545-568
Depuis janvier 2017, la Collection entomologique Ouellet-Robert possède son propre club de curation étudiant, le Club QMOR! Sa mission est de contribuer aux travaux de curation de la collection entomologique Ouellet-Robert tout en faisant la promotion de son importance scientifique et éducative auprès du public. Les objectifs principaux du Club QMOR sont :
1) Accroître, entretenir, et mettre à jour la collection (taxonomie, tiroirs);
2) Échanger des connaissances entomologiques pratiques et théoriques dans une atmosphère sociale.
Le club l’hiver 2018
Les étudiants bénévoles se réunissent à raison de deux soirées par mois au laboratoire d’entomologie du Centre sur la biodiversité (local B-217) de l’Université de Montréal, situé au jardin botanique de Montréal (4101 Sherbrooke Est).
Participer à une rencontre signifie passer une soirée agréable en compagnie d’autres passionnés d’entomologie, tout en travaillant activement à développer notre héritage scientifique. Vous serez appelés à réaliser des projets qui vous permettront de découvrir le fonctionnement d’une collection scientifique, de parfaire vos apprentissages auprès de spécialistes et de développer des compétences avantageuses sur le marché du travail, le tout en ayant la chance d’admirer de superbes insectes indigènes et exotiques!
Les membres inauguraux du club l’hiver 2017, go les «Bibittes anonymes»!
La Collection Ouellet-Robert a été fondée en 1930 par Gustave Chagnon, un des plus grands naturalistes canadiens de son époque. Elle est principalement composée de spécimens capturés par les frères Joseph Ouellet et Adrien Robert. Parmi les plus importantes collections entomologiques du pays, c’est la 1re pour les insectes du Québec.
Intéressés? Nos membres se feront un plaisir de vous accueillir!
Suivez le Club QMOR sur Facebook pour accéder aux photos des réunions précédentes et connaître l’horaire des prochaines.
Classe Arachnida Sous-classe Acari Ordre Trombidiformes Famille Erythraeidae Sous-famille Leptinae GenreLeptusLatreille 1796
Distribution géographique
Les acariens du genre Leptus se retrouvent partout dans le monde, sur tous les continents sauf l’Antarctique (Fain et D’Amico, 1997).
Informations générales et caractères clés
Plusieurs espèces d’acariens ont des larves parasites d’arthropodes. Deux genres parasitent communément les opilions, les genres Trombidium et Leptus. Les larves assez similaires de ces deux genres sont différenciables par la présence d’urstigma (aussi appelés organes de Claparède, de fonction osmorégulatrice) situés entre les deux premières paires de pattes ainsi que la présence d’une ouverture anale chez le genre Trombidium, alors que ces deux caractères sont absents chez le genre Leptus (Krantz, 1978).
Figure II. Larve de Leptus photographiée au microscope à un grossissement de 400x. Notez les soies et les trois paires de pattes. (Spécimen récolté à la Station de biologie des Laurentides en Septembre 2014)
Dans une étude de Welbourn et Young (1988), le genre Leptus était responsable de près de la moitié des cas de parasitisme recensés chez des araignées. Ce genre comprend 90 espèces dont le stade larvaire est parasitaire.
Les mites de Leptus ont une forme plutôt ovale, avec une petite tête portant des chélicères (Faint et Jocqué, 1996). Le céphalothorax et l’abdomen sont fusionnés, comme chez les autres acariens et leur cuticule est molle (Fain et D’Amico, 1997). Les adultes portent quatre paires de pattes, mais la forme larvaire n’en possède que trois (Fain et D’Amico, 1997). Ces mites sont couvertes de soies et la plupart ont une coloration brun-rougeâtre. Elles mesurent d’un à deux milimètres (Bumblebee.org).
Biologie
Leptus, comme plusieurs autres Trombidiformes apparentés, a un cycle de vie particulier avec une alternance de phases de développement actif et de phases inactives, appelées calyptostases, où la stase reste emprisonnée dans la cuticule du stade de développement précédent. Ces acariens comptent six stades de développement post-embryonnaires : la prélarve calyptostasique (inactive), la larve parasitaire, la protonymphe calyptostasique, la deutonymphe prédatrice, la tritonymphe calyptostasique et finalement le stade adulte prédateur, qui est actif (Belozerov, 2008).
Les larves sont parasites de plusieurs ordres différents : des araignées, des opilions, des diptères, des orthoptères et autres. Elles ont des préférences quand à leur site d’attachement à l’hôte : elles choisissent plus souvent la face dorsale, mais certaines espèces de Leptus préfèrent les pattes (Townsend et al, 2006). Ces ectoparasites utilisent leur chélicères pour percer l’arthropode qu’ils parasitent et leur trompe pour en sucer l’hémolymphe. Une substance qui cimente en séchant est aussi sécrétée par des glandes buccales pour solidifier le lien entre l’acarien et son hôte (Abro, 1988). Une fois engorgée, la larve se laisse tomber de l’hôte et se transforme alors en octopode: l’adulte et la deutonymphe sont libres et prédateurs (Pereira et al, 2012).
L’hôte: Phalangium opilio Linnaeus, 1758
Figure III. Phalangium opilio parasité par deux larves : une sur la patte et une sur le coté dorsal (Spécimen récolté à la Station de biologie des Laurentides en Septembre 2014)
Le Phalangium opilio est l’arthropode hôte sur lequel nous avons recueilli nos spécimens de Leptus. Bien que cet opilion commun au Québec ne puisse être utilisé comme un véritable agent de lutte biologique, il est un prédateur utile de plusieurs espèces nuisibles pour l’agriculture, permettant de conserver les densités de ravageurs basses. Bien que cet arachnide soit communément parasité par des mites, on ne connaît pas l’impact du parasitisme sur la survie et la reproduction des opilions (Townsend et al, 2006).
Arnold Åbro (1988) The mode of attachment of mite larvae (Leptus spp.) to harvestmen (Opiliones), Journal of Natural History, 22(1): 123-130
Belozerov, V.N. 2008: Calyptostasy: its role in the development and life histories of the parasitengone mites (Acari: Prostigmata: Parasitengona). Acarina, 16: 3-19.
Fain, A., D’Amico, F., 1997. Observations on Larval Mites (Acari) parasitic on opiliones from the French Pyrenees. Internation Journal of Acarology, 23(1): 39-48
Faint, A., Jocqué, R., 1996. A New Larva of the Genus Leptus Latreille, 1796 (Acari: Erythraeidae) Parasitic on a Spider from Rwanda, International Journal of Acarology, 22(2): 101-108, 1996
Krantz, G. W., 1978. A Manual of Acarology, Second Edition. Oregon State University Book Stores Inc. 509 pages.
Pereira, A.I.A., Fadini, M.A.M., Pikart, T.G., Zanuncio, J.C., Serrao, J.E., 2012. New hosts and parasitism notes for the mite Leptus (Acari: Erythraeidae) in fragments of the Atlantic Forest, Brazil. Brazilian Journal of Biology 72(3): 611-616
Townsend, Jr., V.R., Mulholland, K.A., Bradford, J.O., Proud, D.N., Parent, K.M., 2006. Seasonal variation in parasitism by Leptus mites (Acari, Erythraeidae) upon the harvestman, Leiobunum formosum (Opiliones, Sclerosomatidae). The Journal of Arachnology, 34: 492-494
Welbourn, W.C., Young, O.P., 1988. Mites parasitic on spiders, with a description of a new species of Eutrombidium (Acari: Eutrombidiidae). Journal of Arachnology 16: 373-385
Les espèces de la familles des Syrphidés, aussi connues sous le nom de syrphe, font parties des diptères les plus colorés et les plus fréquemment capturés. Ils se retrouvent sur tous les continents à l’exception de l’Antarctique. Grâce à la grande diversité des niches pouvant être occupées par leurs larves, ils sont présents dans la majorité des types d’habitats, de la forêt tropicale jusqu’aux déserts arides ou arctiques. Étant donné le vaste territoire qu’ils couvrent, ces espèces sont sujettes à une importante pression de la part des prédateurs. C’est pourquoi toutes les espèces de ce groupe montrent une ressemblance générale, et parfois même une ressemblance spécifique très proche avec les abeilles et guêpes (Vockeroth, 1992).
Importance de la famille au niveau économique
Au stade larvaire, la majorité des espèces de syrphidés sont prédatrices d’une grande variété d’espèces appartenant à la super famille des pucerons (Aphidoidea) dont plusieurs espèces sont des ravageurs de cultures. Les syrphidés sont donc d’importants contrôleurs biologiques contre ces insectes (Vokeroth 1992).
Au stade adulte, ils se nourrissent principalement de pollen et de nectar, et représentent donc des pollinisateurs importants pour plusieurs espèces de plantes. En effet ceux-ci transportent le pollen sur leur thorax quand ils butinent aux fleurs (Yariv & Amots, 1976).
Genre Sphaerophoria
Le genre Sphaerophoria regroupe des espèces de mouches de taille petite à médium, ayant des marques jaunes sur la tête, le thorax ainsi que l’abdomen (à noter que certaines espèces sont totalement noires) et leurs corps est plus long que leurs ailes (Wallisroughley, 2014). Il y a 13 espèces connues du genre en Amérique du Nord, distribuées largement au Canada et aux États-Unis.
Espèce Sphaerophoria philanthus
Description
L’espèce mesure environ 10 mm, a un long abdomen à bandes noires et jaunes, le thorax est brun avec une strie brune. L’espèce a des yeux de grande dimension de couleur brune et un visage couleur jaune clair. Les pattes sont jaunes et les ailes transparentes qui comme toutes les espèce de la famille des Syrphidae ont une fausse nervure médiane, la veina spuria. Les femelles se distinguent des mâles par un abdomen se terminant avec un bout plus pointu ainsi des yeux largement séparés sur le front (versus contigus sur le front chez les mâles) (naturesearch 2014).
L’aile présentant la fausse nervure médiane
Les yeux contigus du mâle
Clef de l’espèce
Article apical du tarse des pattes antérieure et médiane brun pâle ou noir et nettement plus foncé que I ‘article basilaire, ou tous les articles de couleur noire. Lobe ventral du surstylus garni d’un appendice pré-apical aplati sur le bord dorsal ; lobe interne du surstylus, pointu dans sa portion apicale (Vokeroth 1992).
Cycle de vie
Sphaerophoria philanthus est une espèce répandue de la famille des Syrphidae à la métamorphose complete, dite holométabole. Quatre stades de métamorphoses caractérisent son cycle de vie : œuf, larve, pupe et adulte (Marshall, 2012).
Stade 1 : Œuf
Les œufs fécondés sont pondus par la femelle à l’aide de la partie terminale télescopique de son abdomen mou et flexible car elle ne possède pas d’appendice ovipositeur (Marshall, 2012 p.16). Les œufs sont déposés sur une plante, près du sol, dès le printemps, tout près ou parmi de leur future source de nourriture, des colonies de pucerons (Coderre, 1987). Ils éclosent rapidement, habituellement en 2 jours (Vockeroth, 1992), une stratégie utilisée lorsque l’habitat est éphémère (Marshall, 2012 p.17)
Stade 2 : Larve
C’est le stade principal d’alimentation. Il correspond à la plus grande partie du cycle de vie de l’insecte. Trois mues larvaires successives et trois instars permettront aux larves de se déplacer, de s’alimenter, de croître et poursuivre leur développement (Marshall, 2012 p.17). Ses larves sont des prédateurs voraces, aphidiphages non spécifiques. Elles se nourrissent de pucerons, hémiptères à cuticule molle, parasites de plantes. Une larve de 3 à 4 jours peut dévorer jusqu’à 300 pucerons par nuit (Agriculture and Agri-Food Canada 2014). Elles les immobilisent à l’aide de sécrétions salivaires collantes et transpercent et aspirent avec leurs pièces buccales verticales en crochet. Elles sont molles, sans capsule céphalique, aveugles, et n’ont pas d’appendice de locomotion. Elles sont colorées jaunes ou vertes grâce à des dépôts de pigments de sang ou de gras sous leur peau translucide. Cette coloration leur permet de se camoufler parmi leur habitat de feuilles et tiges des plantes (Marshall, 2012 p 308).
Stade 3 : Nymphe
Cette étape intermédiaire, permet une première mue de métamorphose (Scott, 2004). Elle est caractérisée par le durcissement de la cuticule du 3e instar larvaire et sa transformation en un cocon, le puparium. La nymphe ne se nourrit pas, son énergie provient des réserves accumulées durant le stade larvaires (Scott, 2004).
Cette transition permettra l’apparition externe des ailes et autres appendices non fonctionnels qui étaient internes chez la larve sous forme de disques imaginaux et la formation de structures de l’adulte (Marshall, 2012 p24). Les variations de températures saisonnières et géographiques durant cette étape, influencent sa coloration. Sphaerophoria philanthus est plus foncé dans les régions plus au sud du Canada (Vockeroth, 1992).
Stade 4 : Adulte
Adulte S. philanthus
Une 2e mue de métamorphose, dite imaginale, permet à l’insecte de compléter son cycle de développement. Des adaptations spécifiques lui permette d’émerger du puparium constitué à cette étape de 3 cuticules soient la cuticule larvaire durcie, celle fine de la nympher, et la cuticule pré-exuviale de l’adulte. À l’aide du ptilinum, une structure gonflable situé sur sa tête et une ligne circulaire plus faible à la partie terminale de son abri il réussit donc à émerger (Marshall, 2012 p.295). Cette nouvelle mouche à fleur pourra s’envoler avec ses deux ailes fonctionnelles, se nourrir non plus de pucerons mais de nectar et de pollen. Les stratégies reproductives peu connues permettront la rencontre d’une femelle et d’un mâle aboutissant à la fécondation des œufs et au recommencement d’un nouveau cycle.
Répartition géographique
La famille des Syrphidae ou mouches à fleurs est très rependue au Canada et aux États-Unis avec ses 870 espèces identifiées. Le genre Sphaerophoria compte 13 espèces en Amérique du Nord (NatureSearch 2014). On les retrouvent en densité élevée dans des secteurs riches en fleurs indigènes avec de larges inflorescences et de petites corolles aplaties structures végétales adaptées à leur morphologie et leur choix alimentaire pour le pollen et le nectar. Dans ces environnements elles ont un rôle essentiel de polinisateur.
On les retrouvent aussi en marges des forêts et des champs cultivées où leurs larves prédatrices aphidiphages trouvent leur source alimentation essentielle et jouent un rôle important dans la lutte biologique contre des parasites de plantes (Sutherland et al 2001) (Martinez et al 2013).
Ecologie
Prédation par les larves
Les larves de Sphaerophoria philanthus se nourrissent d’espèce de la famille Aphidoidea et démontrent l’importance au niveau de la stratégie de différentiation des niches entre les différents prédateurs du groupe. Au niveau de la localisation sur les feuilles d’espèces végétales consommées par les Aphidoidea, l’adulte pond en dessous et seulement sur celles situées à proximité du sol. Au niveau temporel, la ponte à lieu en début de saison (à la mi-juillet) au travers des colonies importantes, notamment de pucerons (Coderre et al. 1987). Bien que les femelles utilisent leur sens visuel pour trouver les lieux convenable pour la ponte, ce serait leur sens de l’odorat qui est le plus important afin de localiser les feuilles occupées par des aphides (Yariv & Amots 1976). Les autres prédateurs ciblent plutôt les feuilles situées plus haut sur la plante et ne s’alimentent pas à la même période de la saison (Coderre et al. 1987).
Mimétisme:
Comme beaucoup d’espèce de la famille des Syrphidae, S. philanthus utilise la stratégie du mimétisme batésien. Elle prend pour modèle la guêpe en ayant une apparence visuelle très ressemblante. Ajoutons qu’elle démontre un comportement semblable, par exemple lorsqu’elle est saisie, elle pousse le bout de l’abdomen vers ce qui la tient, bien qu’elle ne soit pas équipée d’un dard. Un autre exemple est le son émit par la mouche qui rappelle celui des hyménoptères piqueurs (Penney et al 2013).
Coderre, D., and Provencher, L., and Tourneur, J. C. 1987. Oviposition and niche partitioning in aphidophagous insects on maize. Département des Sciences Biologiques, Université du Québec à Montréal, p. 198, 202.
Ce spécimen peut être classé dans l’ordre des Odonata de par le fait qu’il possède des ailes non réduites et qui ne peuvent pas se replier sur le corps. Puisqu’elles sont perpendiculaires au corps au repos et que les postérieures sont plus larges à la base que les antérieures, l’insecte est un anisoptère.
La famille des Aeshnidae est caractérisée par trois éléments morphologiques (2):
-Les triangles formés par les nervures des ailes antérieures et postérieures sont placés à égale distance de l’arculus, soit la séparation de la troisième nervure en plusieurs nervules. De plus, les triangles sont orientés vers l’apex de l’aile de la même façon;
-Le lobe médian du labium est entier;
-Les yeux se touchent sur la ligne médiodorsale.
Les membres du genre Aeshna possèdent un thorax noir ou brun, avec des taches brunes, jaunes ou vertes. Leurs segments abdominaux n’ont qu’une seule carène pour chaque côté. Les Aeshna ont tous une caractéristique particulière au niveau de l’aile: la médiane antérieure et la branche asymétrique qui y est associée forme une fourche qui se dirige vers le bord antérieur de l’aile diagonalement (2).
En général, les membres d’Aeshna sont parmi les plus grands anisoptères vivants. Leur grande taille, ainsi que la coloration particulière de leur corps, sont des caractères suffisants pour une identification préliminaire.
Aeshna umbrosa mesure en moyenne 7,5cm en longueur (1) et leurs ailes mesurent plus que 40mm en longueur. Les membres de l’espèce ont une bande frontoclypéale (soit entre le front et le clypéus, un sclérite qui délimite la marge inférieure de la face) mince et brunâtre, voire de la même couleur que le front et le clypéus.
Cette espèce se distingue des autres Aeshna avec ses deux bandes latérales situées sur le ptérothorax et aucune tache jaune entre les deux bandes. Ces deux bandes latérales sont rectilignes, étroites et de couleur jaune, verte ou bleue, selon le sexe, entourées de noir. Les mâles possèdent des yeux turquoise, ainsi que des bandes dont la couleur varie du bleu-vert au jaune-vert. Les taches abdominales sont très petites et sont de couleur bleue ou verte. Les femelles quant à elles ont des yeux bruns avec des bandes latérales jaune-verdâtre ou des yeux bleutés avec des bandes vert-jaunâtre. Les femelles ont des taches abdominales strictement bleues (3) (4).
Aeshna tuberculifera ressemble beaucoup à Aeshna umbrosa, mais ses bandes ne sont pas entourées de noir et sont plus larges. Le 10e segment abdominal d’A. umbrosa possède des taches ternes, en contraste au 10e segment entièrement noir d’A. tuberculifera. De plus, ce segment possède une saillie épineuse chez A. tuberculifera, alors qu’A. umbrosa en est dépourvu.
On retrouve deux sous-espèces d’Aeshna umbrosa: Aeshna umbrosa umbrosa et Aeshna umbrosa occidentalis. La première possède des taches abdominales vertes plus petites sur les segments postérieures, tandis que la deuxième possède des taches bleues et plus grandes (3).
Habitat
Comme son nom l’indique, l’aeschne des pénombres peut être trouvée dans des endroits avec un peu d’ombre et où l’eau ne bouge pas ou très peu. Cela inclut les lacs, les étangs et les marécages. Cependant, il n’est pas rare de la trouver dans une zone dégagée, surtout lorsqu’elle chasse. C’est dans un tel lieu que nous avons capturé notre spécimen. Pour ce qui est des larves, elles peuvent être trouvées dans les cours d’eau mentionnés précédemment (5).
Diète et techniques de chasse
Toutes les espèces d’odonates sont, sans aucune exception, de redoutables carnivores et ce, au stade larvaire et adulte. Les larves mangent en général les larves des insectes aquatiques, mais peuvent aussi se nourrir de d’autres animaux aquatiques, tels que des crustacés d’eau douce, des tétards et même, des poissons de petite taille. Les adultes quant à eux, se nourrissent de tout insecte ayant une plus petite taille que l’odonate et cela inclut les moustiques, mouches, papillons, coléoptères et même autres odonates. Une particularité de la diète des odonates est qu’ils ne consomment pas les ailes; ils arrachent les ailes de leurs proies avant de les dévorer. On peut dire d’eux qu’ils sont des prédateurs opportunistes (5) (6).
Aeshna umbrosa chasse le plus souvent aux alentours du coucher du soleil, profitant de sa coloration pour se camoufler. En général, les odonates possèdent deux principales techniques de chasse: hawking et gleaming. La première technique consiste en une capture dans les airs d’un autre insecte en vol. La capture se fait principalement avec la bouche, mais peut aussi être complimentée des pattes thoraciques pour bien maintenir la proie. Avec la technique de hawking, Aeshna umbrosa peut consommer quotidiennement jusqu’à 20% de sa masse corporelle (5). La deuxième technique consiste en un survol des zones avec beaucoup de végétation. Lorsque l’odonate aperçoit une proie potentielle, elle plonge vers la proie et l’attrape avec ses pattes thoraciques. Une autre technique peut être utilisée par certaines espèces d’odonates: un individu peut se percher sur un endroit quelconque et attend patiemment le passage d’une proie avant de plonger sur cette dernière (6).
Un odonate en chasse effectue beaucoup de mouvements de va-et-vient afin de patrouiller les environs. Il n’est pas rare d’apercevoir une nuée d’odonates de la même espèce, ou de différentes espèces, qui chassent ensemble. Cette agglomération est due à la présence d’un autre essaim important de potentielles proies dans les parages. Aeshna umbrosa démontre cette caractéristique de former un essaim pour se nourrir (5) (6).
Développement larvaire
Pour réussir à passer à travers l’hiver, les œufs des odonates tombent en diapause, un ralentissement métabolique qui assure leur survie jusqu’au printemps. C’est à ce moment qu’ils commencent à éclore pour libérer des larves (aussi appelées naïades) qui finissent par mesurées entre 3,8 et 4,4 centimètres chez Aeshna umbrosa. Après une période de croissance dans l’eau, elles émergent et passent d’une respiration par branchies anales à une respiration par spiracles thoraciques. Pour compléter leur dernière mue, les larves avalent de l’eau qui les fait gonfler, la cuticule cédant sous la pression qui s’accumule. Les ailes nécessitant une période de sclérification avant de permettre le vol, les odonates sont particulièrement vulnérables à cette période du développement. La mue vers le stade adulte se produit généralement de nuit afin de minimiser la prédation (7).
Distribution
Aeshna umbrosa est répartie à travers le continent nord-américain, dans le sud de la région boréale, de la côte Pacifique jusqu’à la côte Atlantique excluant la province de Terre-Neuve. Cette aeschne se trouve aussi aux États-Unis, allant du Canada jusqu’à la région sud-américaine exclusivement, où le climat sec empêche toute dispersion de l’espèce. La sous-espèce umbrosa est distribuée à l’est du continent, tandis que la sous-espèce occidentalis se trouve à l’ouest (3) (4).
Références
1. « Species Aeshna umbrosa » (en ligne), Iowa State University. 2003-2014. Page consultée le 20 octobre 2014 sur BugGuide.net.
2. Pilon Jean-Guy et Lagacé Denise, 1998. Les odonates du Québec. Entomofaune du Québec (EQ) Inc., 367 pages. p.31, 38-39, 59-64.
3. Paulson Dennis , 2011. Dragonflies and Damselflies of the East. Princeton University Press, 538 pages. p.199-200.
4. Paulson Dennis, 2009. Dragonflies and Damselflies of the West. Princeton University Press, 536 pages. p.222-223.
5. Dunkle Sidney W., 2000. Dragonflies Through Binoculars. Oxford University Press, 369 pages.
Lieu de capture : dans les herbes d’un fossé de drainage, en bordure d’une route en terre battue, à proximité d’une tourbière boisée de la station de biologie des Laurentides. Outil de capture : filet fauchoir Date de capture : 1 septembre 2016 Préparation: d’abord placé dans un récipient de chasse contenant un mélange d’acétate d’éthyle et de plâtre pour le tuer rapidement, avant d’être mis à sécher dans une enveloppe de plastique avec carton
Identification, classification et morphologie
Figure 1 : Front blanc du mâle Sympetrum obtrusum capturé. Photo réalisée par Étienne R.-Dionne.
Frappante, la couleur blanche du front des mâles est l’une des caractéristiques principales permettant de distinguer S. obtrusum parmi les espèces du genre Sympetrum, d’où leur nom vernaculaire “White-faced Meadowhawk”.
Généralement, les individus de cette espèce mesurent de 30 à 35mm. Ils ont les ailes claires. Mâles et femelles présentent un certain dimorphisme. En effet, les femelles n’ont pas le front blanc caractéristique des mâles ni l’abdomen teinté de rouge. Celles-ci, comme les juvéniles, ont plutôt la face d’une couleur qui tire sur le jaune, le brun ou le vert, ainsi qu’un abdomen cendré, ce qui les rend difficiles à distinguer de S. rubicundulum et S. internum (Dunkle, 2000).
Figure 2 : Pièces génitales. A : Appendices anaux supérieurs de l’extrémité de l’abdomen mâle – B : Échancrure de l’hameçon du deuxième segment de l’abdomen mâle – C : Huitième sternite horizontale de l’extrémité de l’abdomen femelle (Ovipositeur) – D: Lames vulvaires du neuvième segment de l’abdomen femelle. Croquis réalisé par Miguelly Bélanger. Inspiré de Robert (1963).
Comme les S. obtrusums, le spécimen capturé possède une face blanche. Cependant, ce sont les appendices anaux supérieurs de l’extrémité de l’abdomen ainsi que l’échancrure de l’hameçon du 2e segment de l’abdomen qui a permis de confirmer son identification (Robert, 1963).
De plus, la position médiane du nodus ainsi que la disposition des nervures principales du supertriangle de l’aile postérieure de notre spécimen sont typiques des Sympetrum (Pilon & Lagacé, 1998). Appartenant également à la famille des libellulidés, leurs ailes postérieures présentent un champ anal en forme de pied possédant un « orteil » plutôt développé. Qui plus est, l’asymétrie de la base des ailes antérieures et postérieures (la base des ailes antérieures étant plus large que celle des ailes postérieures) corrobore l’appartenance au sous-ordre des anisoptères (Silsby, 2001).
Figure 3 : Aile postérieure caractéristique du genre Sympetrum représentant le nodus (A), le pterostigmate (B), le supertriangle et le champ anal. Croquis réalisé par Miguelly Bélanger. Inspiré de Robert (1963).
Dans une perspective d’ensemble, les libellules présentent plusieurs caractéristiques distinctives (Pilon & Lagacé, 1998). Elles ont de courtes antennes. Leurs yeux composés sont de si grandes tailles par rapport au volume de leur tête, qu’ils se rejoignent au sommet de la capsule céphalique. Ils possèdent un long et mince abdomen. Leur thorax orienté vers l’avant permet une capture de proies efficace, en les emprisonnant dans leurs pattes. Au repos, les ailes se posent à plat. Leurs larves possèdent un masque labial étonnant, leur permettant d’atteindre leurs proies avec grande vélocité (Kalkman & al., 2000). Voici un ensemble de caractères partagés par l’espèce du spécimen capturé.
Figure 4 : Taxonomie hiérarchique de Sympetrum obtrusum. Inspiré de Dunkle (2000) et Cresswell & Block (2016).
Les libellules à travers les âges et les continents
Répartis sur l’ensemble de la planète depuis des millénaires, les odonates ont intéressé les hommes et suscité une fascination culturelle à travers les âges (Pilon, 1998). Venant du grec ancien, le mot « odonata » fut créé par Fabricius (1793) pour désigner les « mandibules munies de dents », alors que le mot « Anisoptera » souligne l’asymétrie des ailes (Pilon & Lagacé, 1998). Ceci dit, c’est Linné (1758) qui a introduit le terme « libellules ». En extrême Orient, les libellules sont plutôt bien considérées. Elles y ont d’ailleurs longtemps été utilisées pour traiter les inflammations de la gorge. Au Japon, elles symbolisent la force, la bravoure et la victoire. En Europe, au contraire, ces insectes sont considérés comme maléfiques, une croyance populaire probablement tirée du moyen âge. Pour cette raison, ils se faisaient même appeler « tire-z’yeux » ou « crève-œil » (d’Aguilar, 1985). Plus près d’ici, on a souvent considéré à tort les libellules comme des insectes piqueurs. Aujourd’hui encore, les libellules suscitent un certain engouement.
Évolution
Figure 4 : Représentation des Ères géologiques. Inspirée de Cohen & al. (2013).
Apparus durant la période du carbonifère, les Protodonata sont peut-être les ancêtres des Odonates. Datant du permien, les libellules sont parmi les plus anciens insectes volants (paléoptère) (Grimaldi, D. et Engel, 2005). Dans le même taxon, Protodonata, se trouve le plus grand insecte connu à ce jour (Méganeuropsis permiana), d’une envergure d’ailes de 70cm (Beckemeyer, 2000). Des fossiles de zygoptères et d’anisoptères (Odonata au sens strict) ont d’abord été trouvés au Mésosoïque: les premiers fossiles d’anisoptères modernes datent du Trias (Silsby, 2001). Ceux-ci auraient donc déjà été présents avant que la Pangée se soit complètement disloquée! Émergeant de cette lignée, les libellulidés sont apparus lors de l’Éocène. Avec leurs ailes puissantes et leur corps effilé, leurs ancêtres ont pu bénéficier d’un avantage évolutif important, leur permettant de se distribuer sur un large territoire. À ce jour, on compte près de 7000 espèces d’Odonates (Kalmman & al., 2008) et plus de 60 espèces de Sympetrum à travers le monde (Pilgrim & Dohlen, 2007).
Répartition et habitat
Figure 5 : Carte de la distribution de l’espèce Sympetrum obtrusum. Carte réalisée par Miguelly Bélanger. Inspirée de Dunkle (2000).
Espèce indigène de l’Amérique du Nord, Sympetrum obtrusum se retrouve dans la région zoogéographique Néarctique. Il est principalement recensé au sud du Canada et au nord des États-Unis, mais semble absent du Yukon et du sud de l’Amérique du Nord (Dunkle, 2000; Catling, 2003). Dans ces territoires, il occupe une variété de milieux humides (étangs, lacs, cours d’eau lentiques) et spécialement les tourbières et les marais (Pilon et Lagacé, 1998). Considérant la dispersion des adultes à partir des étendues d’eau, certains avancent que les individus matures constituent une population distincte de celle des larves, malgré le fait que l’un et l’autre occupent les mêmes zones (Faydenko, 2013).
Cycle de vie et alimentation
Essentiels au développement des libellules, les points d’eau douce permettent le développement de leurs œufs et de leurs larves (Kalkman et al., 2008). Généralement, les œufs éclosent environ trois semaines après la ponte, sauf ceux pondus à la fin de la saison estivale, qui tombent en dormance durant l’hiver (Silsby, 2001). C’est d’ailleurs sous forme d’œufs que les embryons de Sympetrum obtrusum passent l’hiver, pour voir leurs larves éclore au printemps et les adultes émerger fin juin, début juillet (Walker & Corbet, 1975). Ainsi, deux générations se succéderont chaque année. Après éclosions, plusieurs stades larvaires s’enchaineront, entrecoupées de mues. Chez les libellules, le premier stade (stade protolarvaire) est caractérisé par l’absence de patte et ne dure qu’une courte période de temps (de quelques secondes à quelques heures) (Silsby, 2001). Par suite, certaines larves de libellules seront plus passives et d’autres plus actives dans leur comportement de chasse, mais toutes sont prédatrices. Grâce à leur masque labial, qui leur permet d’attraper leurs proies à grande vélocité, les larves des Odonates arrivent à se saisir d’insectes, de têtards et même de petits poissons (Corbet, 1999).
Video 1 : Larve de libellule se nourrissant d’une larve de coléoptère à l’aide de son masque labial (par Derek Wheaton, 2015). CC BY-SA 4.0
Contrairement à certains autres genres, le genre Sympetrum consomme non seulement des proies à chacun de ses stades larvaires, mais passe aussi la majeure partie de son cycle de vie sous l’eau (Pritchard, 1964; Corbet, 1999). Les larves de libellules prennent de quelques semaines à quelques années pour se développer, selon l’espèce et différents facteurs environnementaux, tel que la température de l’eau et la quantité de nourriture disponible (Corbet, 1999; Silsby, 2001). Au dernier stade larvaire avant la métamorphose, les derniers organes vitaux finissent de se développer et la larve se prépare pour une dernière mue. L’émergence des libellules matures se fait à l’extérieur de l’eau, sur la végétation ou sur la rive, au courant de la nuit ou au matin, lorsque l’air a pu se réchauffer (Silsby, 2001).
Video 2 : Émergence d’une libellule après métamorphose (auteur inconnu, 2010). CC BY-SA 4.0
Après avoir quitté leur site d’émergence, les libellules occupent la majeure partie de leur temps à chasser, principalement des diptères et de petits insectes volants (Pilon & Lagacé, 1998; Dunkle, 2000). Avec les zygoptères, les libellulidés se distinguent des autres Odonates de par leurLe sympétrum éclaireur démontre une (édité 2018-01-22) mode de chasse à l’affût, repérant leurs proies à partir d’un perchoir, avant de se jeter sur elles (Pilon & Lagacé, 1998). En une seule journée, une libellule peut ainsi consommer jusqu’au cinquième de son poids (Powell & Twist, 1999). Selon les conditions, les libellules en développement auront besoin d’une semaine à un mois pour s’alimenter. Une période cruciale où elles se disperseront afin d’acquérir l’énergie suffisante au développement de leurs organes sexuels (Aguesse,1968; Corbet, 1952). À la recherche de ressources, les mâles Sympetrum obtrusum sont très abondants dans les prairies de Carex. Lors de cette phase de développement, il est possible d’y en observer plus d’un par 10 pi2 (Paulson, 2011) ! Ce n’est qu’en tant qu’individu mature que les mâles et femelles retourneront près de points d’eau pour s’accoupler (Corbet, 1999).
Reproduction
Chez les libellules, le mécanisme de transfert de sperme est indirect (Kalkman, 2008). Après avoir été produit dans les testicules, situés au bout de l’abdomen, le sperme est transféré de manière externe à l’organe copulatoire mâle, à la base de l’abdomen. Grâce à un hameçon, le mâle retient l’abdomen de la femelle durant la copulation. Ainsi, il insémine la femelle et en profite même pour retirer le sperme de ses compétiteurs. Cette compétition spermatique chez les Odonates a été reportée pour la première fois par Waage (1979). Aujourd’hui, c’est ce qui en fait l’un des groupes animaux les plus étudiés pour leur comportement reproducteur (Kalkman, 2008). Suite à la copulation, le mâle reste accroché à la femelle jusqu’à la ponte (Silsby, 2001). Les perforations sur l’une des extrémités des œufs oblongs permettent aux spermatozoïdes de les pénétrer. Puis, les œufs sont ensuite rapidement relâchés à la surface de l’eau ou sur le sol environnant, la femelle ayant un ovipositeur vestigial.
Utilité pour l’homme et statut de conservation
Très sensibles à la qualité de leur habitat et aux perturbations environnementales, les libellules sont souvent utilisées pour évaluer l’ampleur des changements environnementaux, sur le court et le long terme. Aussi, elles sont utilisées comme bioindicateurs de la santé de l’écosystème, dans les projets de conservation et de restauration (Kalman, 2008; Clark & Samways, 1996). Le réchauffement climatique, l’agriculture, l’introduction d’espèce exotique, l’urbanisation et plus encore, entraînent cependant la dégradation et la perte de diversité des milieux humides, dont dépendent les Odonates (Hassall & Thompson, 2008; Kadoya & al., 2009; Rodriguez & al., 2005). De fait, certaines espèces de libellules sont aujourd’hui dans une situation critique (Suh & Samways, 2005; Hämäläinen, 2004). Considérant la large gamme d’organismes interagissant avec les libellules, que ce soit au stade larvaire ou adulte, il est d’autant plus important de protéger les milieux humides, dont ils dépendent.
Bibliographie
Beckemeyer RJ. 2000. The Permian insect fossils of Elmo, Kansas. Emporia State University.
Bulankova E. 1997. Dragonflies (Odonata) as bioindicators of environment quality. Biologia, Bratislava 52:177-180.
Catling PM. 2003. Dragonflies (Odonata) of the Northwest Territories: Status Ranking and Preliminary Atlas. Northwest Territories, Environment and Natural Resources.
Clark TE, Samways MJ. 1996. Dragonflies (Odonata) as indicators of biotope quality in the Kruger National Park, South Africa. Journal of applied ecology:1001-1012.
Cohen K, Finney S, Gibbard P, Fan J-X. 2013. The ICS international chronostratigraphic chart. Episodes 36:199-204.
Corbet PS. 1952. An adult population study of Pyrrhosoma nymphula (Sulzer):(Odonata: Coenagrionidae). The Journal of Animal Ecology:206-222.
Corbet. 1999. Dragonflies: behaviour and ecology of Odonata. Harley Books.
Cresswell S, Block N. 2016. Species Sympetrum – White-faced Meadowhawk. (2016; http://bugguide.net/node/view/8056)
d’Aguilar J, Dommanget J-L. 1985. Guide to the dragonflies of Europe and North Africa. Delachaux et Niestlé.
Dunkle SW. 2000. Dragonflies through binoculars: a field guide to dragonflies of North America. Oxford University Press.
Fabricius JC. 1793. Entomologia systematica emendata et aucta.
Faydenko JL, Butler LM, Swanson BJ. 2013. As the Dragon Flies: Population Structure in Sympetrum obtrusum Dragonflies.
Grimaldi D, Engel MS. 2005. Evolution of the Insects. Cambridge University Press.
Hagen H. 1867. Revision der von Herrn Uhler beschriebenen Odonaten. Stett. Ent. Ztg. 28: 87–95. Sympetrum obtrusum.
Hämäläinen M. 2004. Critical species of Odonata in the Philippines. International Journal of Odonatology 7:305-310.
Hassall C, Thompson DJ. 2008. The effects of environmental warming on Odonata: a review. International Journal of Odonatology 11:131-153.
Kadoya T, Suda S-i, Washitani I. 2009. Dragonfly crisis in Japan: a likely consequence of recent agricultural habitat degradation. Biological Conservation 142:1899-1905.
Kalkman VJ, Clausnitzer V, Dijkstra K-DB, Orr AG, Paulson DR, van Tol J. 2008. Global diversity of dragonflies (Odonata) in freshwater. Hydrobiologia 595:351-363.
Labhart T, Nilsson D-E. 1995. The dorsal eye of the dragonfly Sympetrum: specializations for prey detection against the blue sky. Journal of Comparative Physiology A 176:437-453.
Paulson D. 2011. Dragonflies and Damselflies of the East. Princeton University Press.
Perron J, Jobin L, Mochon A. 2005. Odonatofaune du parc national de la Yamaska, division de recensement de Shefford, Québec. Le Naturaliste canadien 129:17-25.
Pilgrim EM, Von Dohlen CD. 2007. Molecular and morphological study of species-level questions within the dragonfly genus Sympetrum (Odonata: Libellulidae). Annals of the Entomological Society of America 100:688-702.
Pilon J-G, Lagacé D. 1998. Les odonates du Québec: traité faunistique. Entomofaune du Québec.
Powell D, Twist C. 1999. A guide to the dragonflies of Great Britain. Arlequin Press.
Pritchard G. 1964. The prey of dragonfly larvae (Odonata; Anisoptera) in ponds in northern Alberta. Canadian Journal of Zoology 42:785-800.
Robert A. 1963. Les libellules du Québec. Québec: Station biologique du Mont Tremblant, Service de la recherche, Ministère de la chasse et des pêcheries, 1963, 1966 printing.
Rodríguez C, Bécares E, Fernández-Aláez M, Fernández-Aláez C. 2005. Loss of diversity and degradation of wetlands as a result of introducing exotic crayfish. Biological Invasions 7:75-85.
Silsby J. 2001. Dragonflies of the World. Csiro publishing.
Suh AN, Samways MJ. 2005. Significance of temporal changes when designing a reservoir for conservation of dragonfly diversity. Biodiversity & Conservation 14:165-178.
Waage JK. 1979. Dual function of the damselfly penis: sperm removal and transfer. Science 203:916-918.
Walker E, Corbet P. 1975. The Odonata of Canada and Alaska. Vol. 3, Part III: The Anisoptera—Three Families: Toronto: University of Toronto Press.
ATTENTION: Grâce à l’expertise et les commentaires de M. Roch (voir en bas), nous avons mis à jour l’identification de notre spécimen. Il s’agit de Nabis rufusculus et non pas de N. roseipennis comme on avait au paravant. Quelques-unes des phrases suivantes traitent encore de cette deuxième espèce. La biologie des deux espèces est très semblable.
Nabis rugosus – Les espèces du genre Nabis sont généralement uniformes autant dans leurs mode de vie que l’apparence. Michael Becker 2006, CC BY-SA 3.0
Nabis rufusculus est un insecte de l’ordre des hémiptères (punaises), famille des Nabidae. On le retrouve dans une grande partie de l’Amérique du Nord, de la Colombie-Britannique au Colorado à l’Ouest, jusqu’à la Floride et les provinces maritimes à l’Est (Blatchley, 1926). Cette punaise prédatrice passe sa vie dans les herbes au bord de cours d’eau et de marécages, ainsi que sur des arbustes en lisière de forêts.
Historiquement, les espèces de la famille des Nabidae étaient regroupées avec une autre famille d’hémiptères, les Reduviidae, qui sont souvent nommés « punaises assassines ». Les réduves ont une apparence mais aussi un mode de vie très similaire aux Nabidae, et c’est en 1861 que Nabidae a obtenu le statut de famille (Schuh et al., 1995). Le genre Nabis a quant à lui été décrit par Latreille bien avant cette séparation, soit en 1802, et l’espèce N. rufusculus en 1872 par Reuter.
Comme tous les hémiptères, N. rufusculus a des pièces buccales modifiées en forme de trompe, nommée rostre (Dolling, 1991). Il est articulé en sections, contrairement au proboscis, la trompe des lépidoptères (papillons) entièrement flexible. Le nombre de sections du rostre, 4 chez les Nabidae, est un des critères permettant de différencier avec les Reduviidae, qui compte seulement 3 articles (Borror et al., 1991). Le rostre est un appareil polyvalent qui sert à piquer les tissus des plantes ou des animaux pour en aspirer les substances nutritives, ce qui a contribué à la diversité des modes de vie chez les Hémiptères.
Patte ravisseuse- noter les crochets sur l’intérieur du tibia
Nabis rufusculus est quant à lui un prédateur dit généraliste, capable de se nourrir d’une grande variété de proies, à un point tel que l’espèce adopte certains comportements pour réduire le cannibalisme. Ainsi, la larve est généralement située plus bas sur une même plante que l’adulte, probablement pour diminuer les chances de rencontres (Braman et al., 1989). En conditions de laboratoire, les larves doivent aussi être gardées séparément pour éviter qu’elles ne se mangent entre elles (Rensner et al., 1983). Outre ses propres congénères, les adultes et nymphes mangent une variété d’insectes à cuticule molle comme les pucerons, ainsi que des larves, des nymphes ou des œufs d’autres espèces (Henry et al., 1988). Les punaises prédatrices capturent généralement leurs proies à l’aide de leurs pattes dites “ravisseuses”. Comme celles des mantes religieuses, leur tibia et leur fémur des pattes prothoraciques (les pattes avant) sont dotés de crochets ou de poils pour retenir la proie entre ces deux segments de la patte (Dolling, 1991). Elles piquent ensuite avec leur rostre, injectent des enzymes pour digérer la proie de l’intérieur, pour ensuite aspirer le contenu avec ce même rostre. En conditions de laboratoire, Rensner (1983) a observé que N. roseipennis consommait en moyenne 4 à 5 cicadelles de la pomme de terre (Empoasca fabae) adultes en 24h; elle contribue donc à contrôler les populations de ce ravageur de plusieurs grandes cultures. Les Nabidae sont des prédateurs importants d’autres ravageurs, dont le ver de l’épi de maïs, Helicoverpa zea (Swenson et al, 2013), la teigne des crucifères, Plutella xylostella (Philips et al., 2014), et plusieurs espèces de pucerons (Chasen et al. 2014). Quelques cas de piqûres ont été répertoriés chez l’humain par des espèces du genre Nabis (Faúndez et Carvajal, 2011), mais pas N. roseipennis. Contrairement à d’autres familles de punaises qui se nourrissent de sang, les Nabidae ne s’en prennent généralement pas aux vertébrés. Enfin, les Nabidae piquent parfois les végétaux pour s’y abreuver et compléter leur diète, lorsque les proies sont rares (Stoner A., 1972).
Clasper de Nabis rufusculus
Lors de la reproduction, le mâle approche la femelle en émettant des stridulations similaires à celles d’un grillon, en frottant les tibias de ses pattes mésothoraciques (pattes du milieu). Au contact de la femelle, le mâle utilise ses claspers, deux structures situées de part et d’autre de l’abdomen du mâle. Similaire à des pinces, les claspers servent à retenir la femelle durant l’accouplement (Davies, 1988). C’est d’ailleurs la forme des claspers d’un individu, parfois asymétriques, qui est souvent le meilleur critère pour l’identification de nombreuses espèces de punaises, dont N. rufusculus qui peut être facilement confondue avec d’autre espèce du genre Nabis à l’allure très similaire (Larochelle, 1984). Les claspers plus large, en forme de fer de hache, distinguent ainsi N. roseipennis de N. rufusculus, dont les claspers sont plus étroits. [Voir commentaires en bas]
La femelle pond des œufs d’à peine 2 mm dans la tige des végétaux, avec seulement l’opercule servant d’écoutille visible à l’extérieur de la plante (Dolling, 1991). À l’éclosion, la larve pousse l’opercule et s’extrait de l’œuf avant d’effectuer une pause près du trou d’émergence. Cette pause permet à sa cuticule encore neuve de sécher et durcir. La larve ressemble beaucoup à un petit adulte sans ailes. Il y a cinq stades larvaux avant l’âge adulte, donc quatre mues successives au cours desquelles les futures ailes se développent avant la mue finale. Nabis rufusculus semble avoir une préférence pour les légumineuses comme site de ponte, du moins par rapport à d’autres grandes cultures; elle est donc fréquemment observée dans les champs de soja et de luzerne (Pfannenstiel et al., 1998) où les proies sont également abondantes. Il y a généralement deux pontes dans l’été sous notre climat québécois, après quoi les adultes, comme tous les Nabis, hibernent sous une litière de feuilles mortes (Dolling, 1991).
Référence
Blatchley, W. S. (1926) Heteroptera, or true bugs of eastern North America, with especial reference to the faunas of Indiana and Florida. The Nature publishing company, Indianapolis, 1136p.
Borror, D. J., White, R. E. (1991) Le guide des insectes du Québec et de l’Amérique du Nord. Broquet, Boucherville, 408p.
Henry, T. J., Froeschner, R. C. (1988) Catalog of the Heteroptera, or true bugs, of Canada and the continental United States. E. J. Brill and Leiden, New York, 958p.
Larochelle, A. (1984) Les punaises terrestres (Heteroptères: Geocorises) du Québec, Association des entomologistes amateurs du Québec. Québec, 513p.
Schuh, R. T., Slater, J. A. (1995) True bugs of the world (Hemiptera: Heteroptera) Classification and natural history. Cornell University Press, New York, 336p.














































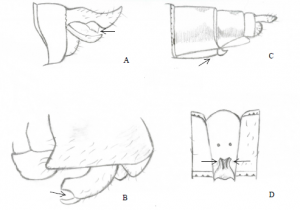









{kind=link}
{kind=link}